Neurotransmitterafgifte
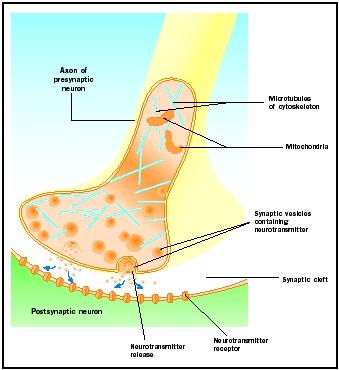
Actiepotentialen die bij synaptische knoppen aankomen, brengen de afgifte van neurotransmitter in de synaptische spleet op gang. Het moleculaire mechanisme wordt niet volledig begrepen. Tussen de aankomst van de actiepotentiaal en de afgifte van neurotransmitter treedt een “synaptische vertraging” op van één tot twee milliseconden. Actiepotentialen openen calciumkanalen in het membraan van de synaptische knop, waardoor een binnenwaartse beweging van calciumionen ontstaat. Calciumionen triggeren het vrijkomen van neurotransmitter uit synaptische blaasjes in de synaptische spleet. De synaptische blaasjes versmelten met het presynaptische membraan tijdens dit proces van exocytose. De membranen van oude blaasjes gaan deel uitmaken van de presynaptische membraan en nieuwe blaasjes knellen af van een aangrenzend gebied van de membraan. Deze nieuwe blaasjes worden vervolgens weer gevuld met nieuw gesynthetiseerde of “gerecycleerde” neurotransmitters .
De vrijgekomen neurotransmitters verspreiden zich over de smalle synaptische spleet. Op het postsynaptische membraan binden neurotransmittermoleculen zich aan membraangebonden receptormoleculen met herkenningsplaatsen die specifiek zijn voor die neurotransmitter. Binding van de neurotransmitter aan de receptor leidt tot een postsynaptische respons die specifiek is voor die receptor. Deze reacties kunnen excitatoir of inhibitoir zijn, afhankelijk van de eigenschappen van de receptor. Als stimulatie van de receptor ertoe leidt dat het postsynaptische membraan elektrisch positiever wordt (depolariseert), is er sprake van een excitatoir postsynaptisch potentiaal (EPSP). Als het membraan negatiever wordt (hypergepolariseerd), is er sprake van een inhibitoire postsynaptische potentiaal (IPSP). Excitatie en inhibitie hangen af van de eigenschappen van de receptor en niet van de neurotransmitter. Receptoren gekoppeld aan natrium- of calciumkanalen zijn excitatoir en veroorzaken een depolarisatie van het postsynaptische membraan, terwijl receptoren gekoppeld aan chloride- of kaliumkanalen inhibitoir zijn en een hyperpolarisatie van het postsynaptische membraan veroorzaken. Dergelijke receptoren gekoppeld aan ionkanalen worden ionotrope receptoren genoemd.
Andere receptoren zijn gekoppeld aan “second-messenger” systemen die een reeks biochemische reacties in de postsynaptische cel in gang zetten. Dit zijn metabotrope receptoren. Metabotrope receptoren kunnen veel verschillende
postsynaptische gebeurtenissen. Deze variëren van de directe activering van aangrenzende ionenkanalen, tot wijziging van de receptorgevoeligheid, tot transcriptie van specifieke boodschapper-ribonucleïnezuren (RNA’s), of zelfs de activering van specifieke genen . Chemische synapsen maken deel uit van een zeer aanpasbaar en flexibel communicatiesysteem. Het zijn geen statische anatomische structuren met vaste eigenschappen, maar het zijn dynamische structuren, die in staat zijn hun moleculaire eigenschappen te veranderen bij veranderende omstandigheden.
Er zijn letterlijk honderden neurotransmitters. Sommige zijn vrij eenvoudige verbindingen zoals acetylcholine, serotonine, de catecholamines (dopamine, noradrenaline, en epinefrine) en een aantal van de aminozuren . Vele zijn complexer en behoren tot de grote reeks neuropeptide transmitters. Eenmaal in de synaptische spleet vrijgemaakt, blijven neurotransmitters actief totdat zij chemisch veranderd zijn of door speciale transportsystemen weer in de synaptische knop worden opgenomen en gerecycleerd. Bij cholinerge synapsen is acetylcholinesterase aanwezig in de synaptische spleet. Dit enzym splitst de neurotransmitter in acetaat en choline, die geen van beide actief zijn. Serotonine en epinefrine daarentegen worden opgenomen in de presynaptische terminal en gerecycleerd.
Geef een antwoord