Subphylum Crustacea
General Nature of a Crustacea
Crustacea verschillen op verschillende manieren van andere geleedpotigen, maar het enige echt onderscheidende kenmerk is dat crustacea de enige geleedpotigen zijn met twee paar antennes. Naast twee paar antennes en een paar onderkaken hebben kreeftachtigen twee paar maxillae op de kop, gevolgd door een paar aanhangsels op elk lichaamssegment of somiet. Bij sommige kreeftachtigen zijn niet alle somieten voorzien van aanhangsels. Alle aanhangsels, behalve misschien de eerste antennes, zijn primitief birameeus (twee hoofdtakken), en althans sommige aanhangsels van de huidige volwassenen vertonen die toestand. Organen gespecialiseerd in ademhaling, indien aanwezig, functioneren als kieuwen.
De meeste kreeftachtigen hebben tussen 16 en 20 somieten, maar sommige vormen hebben 60 somieten of meer. Een groter aantal somieten is een primitief kenmerk. De meer afgeleide toestand is het hebben van minder segmenten en een grotere tagmatisatie. De belangrijkste tagmata zijn kop, thorax en abdomen, maar deze zijn niet in de hele klasse (of zelfs binnen sommige subklassen) homoloog vanwege de verschillende mate van samensmelting van somieten, bijvoorbeeld zoals bij de cephalothorax.
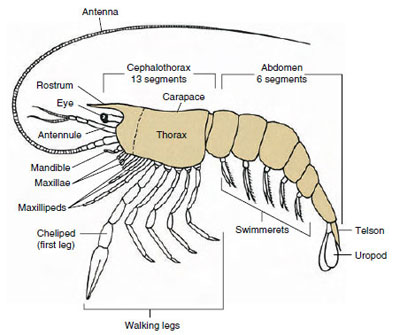
Figuur 19-1
Archetypische plattegrond van Malacostraca. De twee maxillae en drie
maxillipedia zijn schematisch van elkaar gescheiden om het
algemene plan te illustreren.
Verreweg de grootste groep kreeftachtigen is de klasse Malacostraca, die kreeften, krabben, garnalen, strandspringers, zeugwantsen, en vele andere omvat. Deze vertonen een verrassend constante schikking van lichaamssegmenten en tagmata, die wordt beschouwd als het voorouderlijke plan van de klasse (figuur 19-1). Dit typische lichaamsplan heeft een kop van vijf (zes embryonaal) vergroeide somieten, een borststuk van acht somieten, en een achterlijf van zes somieten (zeven bij enkele soorten). Aan het vooreinde bevindt zich een niet gesegmenteerd rostrum en aan het achtereind een niet gesegmenteerd telson, dat met de laatste buiksomiet en zijn uropoden bij vele vormen een staartwaaier vormt.
Bij vele kreeftachtigen kan het rugschild van de kop zich naar achteren en rond de zijkanten van het dier uitstrekken om sommige of alle thoracale en buiksomieten te bedekken of ermee vergroeid te zijn. Deze bedekking wordt een carapax genoemd. Bij sommige groepen vormt het kopborststuk schelpvormige kleppen die het gehele lichaam of een groot deel daarvan bedekken. Bij de decapoden (o.a. kreeften, garnalen en krabben) bedekt het kopborststuk de hele cephalothorax, maar niet het achterlijf.
Vorm en functie
Omwille van hun grootte en gemakkelijke verkrijgbaarheid zijn grote kreeftachtigen zoals rivierkreeften meer bestudeerd dan andere groepen. Zij worden ook vaak bestudeerd in inleidende laboratoriumcursussen. Daarom zijn veel van de hierna volgende opmerkingen specifiek van toepassing op kreeften en hun verwanten.
Uiterlijke kenmerken
De lichamen van kreeftachtigen zijn bedekt met een afgescheiden cuticula die bestaat uit chitine, eiwit en kalkhoudend materiaal. De hardere, zware platen van grotere kreeftachtigen zijn bijzonder rijk aan kalkhoudende afzettingen. De harde beschermende bekleding is zacht en dun bij de gewrichten tussen de somieten, waardoor een soepele beweging mogelijk is. Het kopborststuk, indien aanwezig, bedekt een groot deel of het gehele kopborststuk; bij decapoden zoals kreeften worden alle kop- en borstsegmenten dorsaal omsloten door het kopborststuk. Elke somiet die niet door het kopborststuk wordt omsloten, is bedekt met een dorsale cuticulaire plaat, of tergum (figuur 19-2A), en een ventrale dwarsstaaf, het sternum, ligt tussen de segmentale aanhangsels (figuur 19-2B). Het achterlijf eindigt in een telson, dat niet als een somiet wordt beschouwd en de anus draagt. (Het telson is mogelijk homoloog met het pygidium van de eenpotigen.)
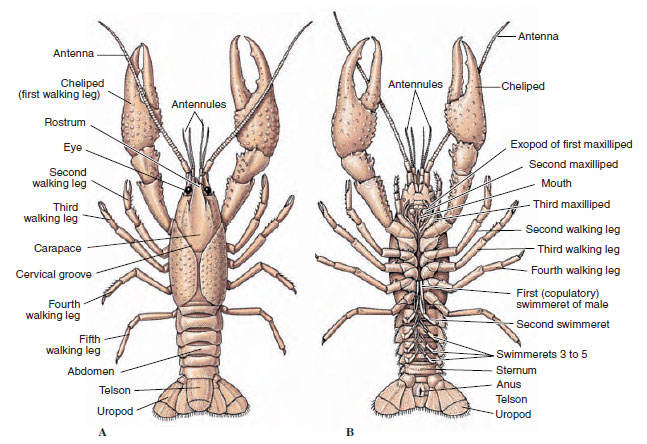
Figuur 19-2
Uitwendige structuur van rivierkreeften. A, Dorsaal zicht. B, Ventraal aanzicht.
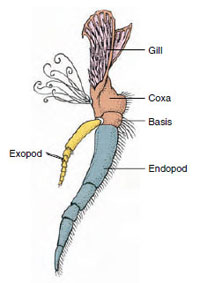
Figuur 19-3
Delen van een biaamvormig schaaldier
aanhangsel (derde maxillipedum van een
kreeftje).
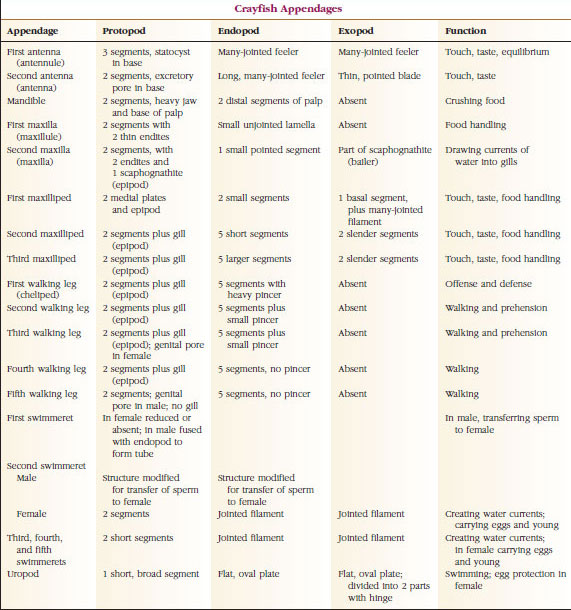
De plaats van de gonoporen varieert naar gelang van het geslacht en de groep kreeftachtigen. Zij kunnen zich bevinden op of aan de basis van een paar aanhangsels, aan het uiteinde van het lichaam, of op somieten zonder poten. Bij kreeften bevinden de openingen van de vasa deferentia zich aan de mediaanzijde aan de basis van het vijfde paar wandelpoten, en die van de eileiders aan de basis van het derde paar. Bij de wijfjes bevindt de opening naar de zaadleider zich meestal in de midventrale lijn tussen het vierde en vijfde paar loopbenen.
Appendages: De leden van de klassen Malacostraca (met inbegrip van de kreeften) en Remipedia hebben gewoonlijk een paar vergroeide aanhangsels op elke somiet (Figuur 19-3), hoewel de buiksomieten in de andere klassen geen aanhangsels dragen. Aanzienlijke specialisatie is duidelijk bij aanhangsels van afgeleide schaaldieren zoals kreeften. Het zijn echter allemaal variaties van het biramide basisplan, geïllustreerd door een aanhangsel van een rivierkreeft zoals een maxilliped (een thoracaal ledemaat dat is omgevormd tot een hoofdaanhangsel) (Figuren 19-3 en 19-4). Het basale gedeelte, of protopode, draagt een laterale exopode en een mediale endopode. De protopode bestaat uit één of twee gewrichten (basis en coxa), terwijl de exopode en de endopode elk één tot verscheidene gewrichten hebben. Sommige aanhangsels, zoals de wandelpoten van kreeften, zijn secundair uniramidaal geworden. Soms zijn er mediale of laterale uitsteeksels op de ledematen van schaaldieren, die respectievelijk endites en exites worden genoemd, en een exite op de protopode wordt een epipode genoemd. Epipoden zijn vaak omgevormd tot kieuwen. Tabel 19-1 laat zien hoe de verschillende aanhangsels van het biramous plan zijn gewijzigd om specifieke functies te vervullen.
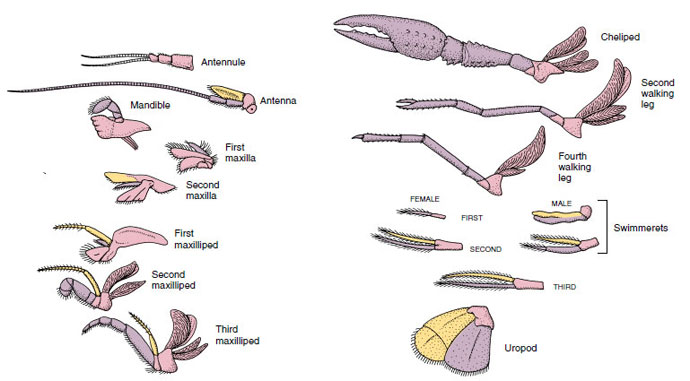
Figuur 19-4
Aanhangsels van een rivierkreeft die laten zien hoe ze zijn gemodificeerd ten opzichte van het biramous basisplan, zoals aangetroffen in een
zwemmertje. Protopode, roze; endopode, paars; exopode, geel.
Structuren die een soortgelijk basisplan hebben en van een gemeenschappelijke vorm zijn afgekalfd, worden homoloog genoemd, of ze nu dezelfde functie hebben of niet. Aangezien gespecialiseerde loopbenen, monddelen, kaakslippen en zwemvliezen zich alle hebben ontwikkeld uit een gemeenschappelijk biramateus type, maar zijn aangepast om verschillende functies te vervullen, zijn zij alle homoloog aan elkaar, een toestand die bekend staat als seriële homologie. Primitief waren de ledematen allemaal zeer gelijksoortig, maar tijdens de evolutie van de structurele wijzigingen zijn sommige takken verkleind, sommige verloren gegaan, sommige sterk gewijzigd, en sommige nieuwe delen toegevoegd. Rivierkreeften en hun verwanten bezitten de meest uitgebreide seriële homologie in het dierenrijk, met 17 verschillende maar serieel homologe types aanhangsels (Tabel 19-1).
TABEL 19.1
Interne kenmerken
Het spier- en zenuwstelsel en de segmentatie in de thorax en het abdomen vertonen duidelijk metamerisme dat van de annelide-achtige voorouders is geërfd, maar er zijn duidelijke wijzigingen in andere systemen. De meeste veranderingen betreffen concentratie van delen in een bepaalde regio of anders reductie of volledig verlies van delen, zoals de intersepta.
Hemocoel: De belangrijkste lichaamsruimte bij geleedpotigen is niet het coëloom maar een met bloed gevulde hemocoel. Tijdens de embryonale ontwikkeling van de meeste geleedpotigen openen zich in het mesoderm van ten minste enkele somieten rudimentaire coelomische holten. Deze worden spoedig uitgewist of gaan over in de ruimte tussen de zich ontwikkelende mesodermale en ectodermale structuren en de dooier. Deze ruimte wordt de hemocoel en is dus niet bekleed met een mesodermaal peritoneum. Bij schaaldieren zijn de enige coelomische compartimenten die overblijven de eindzakken van de uitscheidingsorganen en de ruimte rond de gonaden.
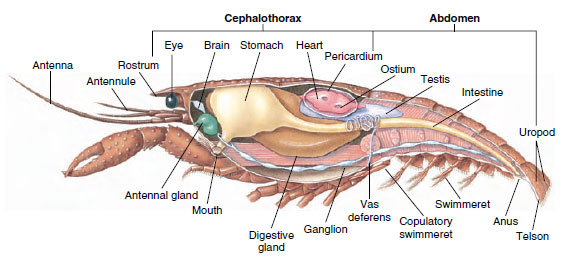
Musculair systeem: Gestreepte spieren maken een aanzienlijk deel uit van het lichaam van de meeste Crustacea. De spieren zijn gewoonlijk gerangschikt in tegengestelde groepen: buigers, die een deel naar het lichaam toe trekken, en strekkers, die het naar buiten uitstrekken. Het achterlijf van een rivierkreeft heeft krachtige buigspieren (Figuur 19-5), die worden gebruikt wanneer het dier achteruit zwemt – zijn beste manier om te ontsnappen. Sterke spieren aan weerszijden van de maag controleren de onderkaken.
Figuur 19-5
Inwendige structuur van een mannelijke rivierkreeft.
Respiratoir systeem: Bij kleinere schaaldieren vindt de gasuitwisseling plaats over dunnere delen van de cuticula (bijvoorbeeld in de poten) of over het hele lichaam, en gespecialiseerde structuren kunnen afwezig zijn. Grotere kreeftachtigen hebben kieuwen, die delicate, veerachtige uitsteeksels zijn met een zeer dunne cuticula. Bij decapoden omsluiten de zijkanten van het kopborststuk de kieuwholte, die anterior en ventraal open is (figuur 19-6). De kieuwen kunnen uitsteken vanuit de borstwand in de kieuwholte, vanuit de verbinding van de thoracale poten met het lichaam, of vanuit de thoracale coxae. De laatste twee types zijn typisch voor kreeften. De “puls”, een deel van de tweede bovenkaak, trekt water over de kieuwdraden, in de kieuwholte aan de bases van de poten, en uit de kieuwholte aan de voorste.
Circulatoir systeem: Schaaldieren en andere geleedpotigen hebben een “open” of lacunair systeem van bloedsomloop. Dit betekent dat er geen aders zijn en geen scheiding van bloed en interstitiële vloeistof, zoals bij dieren met een gesloten systeem. Hemolymfe (bloed) verlaat het hart via slagaders, circuleert door de hemocoel, en keert terug naar veneuze sinussen, of ruimten, in plaats van aders, voordat het weer het hart ingaat. Anneliden hebben een gesloten systeem, evenals gewervelde dieren.
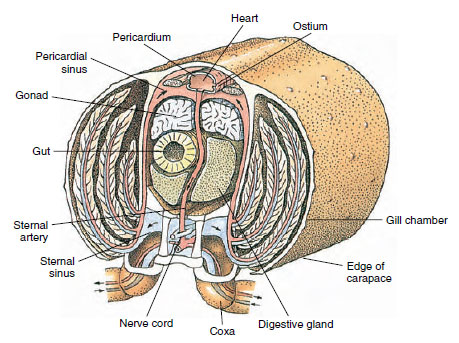
Een dorsaal hart is het voornaamste voortstuwingsorgaan. Het is een eenkamerige zak van gestreepte spieren. Hemolymfe komt het hart binnen vanuit de omringende sinus pericardiale via gepaarde ostia, met kleppen die terugstroming naar de sinus voorkomen (figuur 19-6). Vanuit het hart komt hemolymfe in een of meer slagaders. Kleppen in de slagaders voorkomen een terugstroom van hemolymfe. Kleine slagaders monden uit in weefselsinus, die op zijn beurt vaak uitmondt in een grote sternale sinus (figuur 19-6).
Van daaruit voeren afferente sinuskanalen hemolymfe naar de kieuwen, indien aanwezig, voor de uitwisseling van zuurstof en kooldioxide. Hemolymfe keert vervolgens via efferente kanalen terug naar de sinus pericardialis (figuur 19-6).
Figuur 19-6
Diagrammatische dwarsdoorsnede door het hartgebied van een rivierkreeft, waarop de richting van de bloedstroom in dit “open”
bloedsysteem is te zien. Het hart pompt bloed naar de lichaamsweefsels via slagaders, die uitmonden in weefsel sinussen. Terugkerend
bloed komt in de sternale sinus, gaat dan door de kieuwen voor gasuitwisseling, en tenslotte terug naar de pericardiale sinus via efferente
kanalen. Let op de afwezigheid van aderen.
Hemolymfe bij geleedpotigen is grotendeels kleurloos. Het bevat ameboide cellen van ten minste twee typen. Hemocyanine, een koperhoudend ademhalingspigment, of hemoglobine, een ijzerhoudend pigment, kunnen in oplossing worden meegevoerd. Hemolymfe heeft de eigenschap te stollen, wat het verlies ervan bij kleine verwondingen voorkomt. Sommige ameboïde cellen geven een trombineachtige stollingsstof af die de stolling neerslaat.
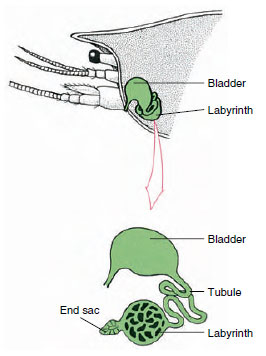
Figuur 19-7
Schema van de antennaklier (groene klier) van
kreeftjes. (In natuurlijke positie is het orgaan veel
gevouwen.) Sommige schaaldieren hebben geen labyrint,
en de uitscheidingsbuis (nefridiaal kanaal) is
een sterk opgerolde buis.
Uitscheidingsstelsel: De uitscheidingsorganen van volwassen schaaldieren zijn een paar buisvormige structuren die zich in het ventrale deel van de kop bevinden, vóór de slokdarm (figuur 19-5). Ze worden antenneklieren of maxillaire klieren genoemd, afhankelijk van de vraag of ze opengaan aan de basis van de antennes of van de tweede maxillae. Enkele volwassen kreeftachtigen hebben beide. De uitscheidingsorganen van decapoden zijn antennaklieren, die in deze groep ook wel groene klieren worden genoemd. Schaaldieren hebben geen malpighian tubules, de uitscheidingsorganen van spinnen en insecten.
De eindzak van de antennaalklier, die is afgeleid van een embryonaal coelomisch compartiment, bestaat uit een klein blaasje (saccule) en een sponsachtige massa die labyrint wordt genoemd. Het labyrint staat via een uitscheidingsbuisje in verbinding met een dorsale blaas, die naar buiten opengaat via een porie op het ventrale oppervlak van het basale antennale segment (Figuur 19-7). De hydrostatische druk in de hemocoel oefent kracht uit voor de filtratie van vloeistof in de eindzak. Terwijl het filtraat door de uitscheidingsbuis en de blaas gaat, wordt het gewijzigd door resorptie van zouten, aminozuren, glucose en wat water, en uiteindelijk uitgescheiden als urine.
Excretie van stikstofhoudende afvalstoffen (vooral ammoniak) vindt plaats door diffusie over dunne oppervlakken van de cuticula, vooral de kieuwen, en de zogenaamde uitscheidingsorganen functioneren voornamelijk om de ionische en osmotische samenstelling van lichaamsvloeistoffen te regelen. Zoetwaterschaaldieren, zoals rivierkreeften, worden voortdurend bedreigd door overdilutie van hun bloed door water, dat over de kieuwen en andere waterdoorlatende oppervlakken diffundeert. De groene klieren, die een verdunde, zoutarme urine vormen, fungeren als een doeltreffend “overstromingscontrole”-apparaat. Er gaat wat Na+ en Cl- verloren in de urine, maar dit verlies wordt gecompenseerd door actieve absorptie van opgelost zout door de kieuwen. Bij zeewaterschaaldieren, zoals kreeften en krabben, regelt de nier de zoutsamenstelling van de hemolymfe door selectieve wijziging van het zoutgehalte van de urinebuisjes. Bij deze vormen blijft de urine isosmotisch ten opzichte van het bloed.
Nervus- en zintuigstelsels: De zenuwstelsels van schaaldieren en anneliden hebben veel gemeen, hoewel die van schaaldieren meer fusie van ganglia vertonen (figuur 19-5). De hersenen zijn een paar supraesofageale ganglia die zenuwen leveren aan de ogen en twee paar antennes. Het is verbonden door bindweefsels met het subesofageale ganglion, een fusie van ten minste vijf paar ganglia die zenuwen leveren aan de mond, de aanhangsels, de slokdarm en de antenneklieren. Het dubbele ventrale zenuwkoord heeft een paar ganglia voor elke somiet en zenuwen die de aanhangsels, spieren en andere delen bedienen.
Naast dit centrale systeem kan er een sympathisch zenuwstelsel zijn dat verbonden is met het spijsverteringskanaal.
Schaaldieren hebben beter ontwikkelde zintuigen dan anneliden. De grootste zintuigen van kreeften zijn de ogen en de statocysten. Wijd verspreid over het lichaam zijn tastharen, delicate uitsteeksels van cuticula die vooral overvloedig voorkomen op chelae, monddelen, en telson. De chemische zintuigen van smaak en reuk worden gevonden in haren op antennes, monddelen, en andere plaatsen.
Een zakachtige statocyst, die naar de oppervlakte opent door een dorsale porie, wordt gevonden op het basale segment van elke eerste antenne van kreeften. De statocyst bevat een richel waarop sensorharen zitten die gevormd zijn uit de chitineuze bekleding en zandkorrels die dienen als statolieten. Telkens wanneer het dier zijn positie verandert, worden overeenkomstige veranderingen in de positie van de korrels op de sensorharen als prikkels doorgegeven aan de hersenen, en kan het dier zich dienovereenkomstig aanpassen. Elke vervelling (ecdysis) van de cuticula resulteert in het verlies van de cuticulaire bekleding van de statocyst en daarmee van de zandkorrels. Nieuwe korrels worden na de ecdysis door de dorsale porie opgenomen.
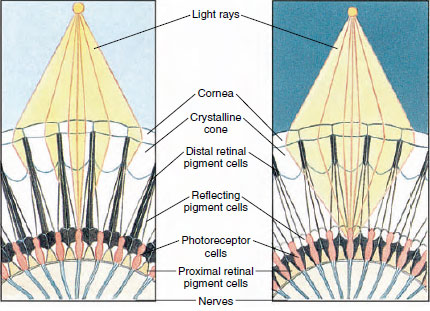
Figuur 19-8
Deel van het samengestelde oog van een geleedpotige waarop de migratie van pigment
in ommatidia voor dag- en nachtzicht te zien is. In
elk diagram zijn vijf ommatidia afgebeeld. Overdag is elk ommatidium omgeven door een donkere
pigmentkraag zodat elk ommatidium alleen wordt gestimuleerd door lichtstralen
die zijn eigen hoornvlies binnendringen (mozaïsch zicht); ’s nachts vormt het pigment
onvolledige kragen en kunnen de lichtstralen zich verspreiden naar aangrenzende ommatidia
(continu, of superpositie, beeld).
De ogen van veel schaaldieren zijn samengesteld en bestaan uit vele fotoreceptoreceptoreenheden, ommatidia genaamd (figuur 19-8). Het afgeronde oppervlak van elk oog wordt bedekt door een doorzichtig stuk cuticula, het hoornvlies, dat verdeeld is in vele kleine vierkanten of zeshoeken, facetten genoemd. Deze facetten zijn de buitenste uiteinden van de ommatidia. Elk ommatidium gedraagt zich als een klein oog en bevat verschillende soorten cellen die op zuilvormige wijze zijn gerangschikt (Figuur 19-8). Tussen de aangrenzende ommatidia bevinden zich zwarte pigmentcellen.
De beweging van het pigment in een samengesteld oog van een geleedpotige stelt hem in staat zich aan te passen aan verschillende hoeveelheden licht. Er zijn drie reeksen pigmentcellen in elk ommatidium: distale retinale, proximale retinale, en reflecterende; deze zijn zo gerangschikt dat ze een min of meer volledige kraag of huls rond elk ommatidium kunnen vormen. Bij sterk licht of dagadaptatie beweegt het distale retinale pigment naar binnen en ontmoet het naar buiten bewegende proximale retinale pigment, zodat zich rond het ommatidium een volledige pigmentkoker vormt (figuur 19-8). In deze toestand zullen alleen de stralen die het hoornvlies rechtstreeks treffen, de fotoreceptorcellen (retinula) bereiken, want elk ommatidium is afgeschermd van de andere. Elk ommatidium ziet dus slechts een beperkt deel van het gezichtsveld (een mozaïek- of appositiebeeld). Bij zwak licht scheiden distale en proximale pigmenten zich, zodat de lichtstralen, met behulp van reflecterende pigmentcellen, de kans krijgen zich te verspreiden naar aangrenzende ommatidia en een continu, of superpositie, beeld te vormen. Dit tweede type zicht is minder nauwkeurig, maar maakt maximaal gebruik van de beperkte hoeveelheid ontvangen licht.
Reproductie, levenscycli en endocriene functie
De meeste kreeftachtigen hebben gescheiden geslachten, en er zijn verschillende specialisaties voor copulatie bij verschillende groepen. Zeepokken zijn eenhuizig maar doen over het algemeen aan kruisbevruchting. Bij sommige ostracoden zijn mannetjes schaars, en is de voortplanting gewoonlijk parthenogenetisch. De meeste kreeftachtigen broeden hun eieren op een of andere manier: branchiopoden en zeepokken hebben speciale broedkamers, roeipootkreeftjes hebben broedzakken die aan de zijkanten van het achterlijf zijn bevestigd (zie figuur 19-19), en veel malacostracans dragen eieren en jongen die aan hun abdominale aanhangsels zijn bevestigd.
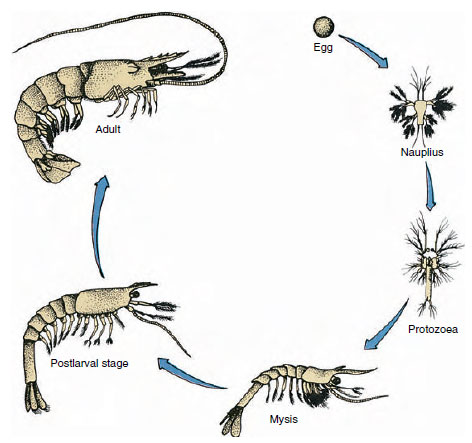
Figuur 19-9
Life cycle of a Gulf shrimp Penaeus. Penaeiden paaien op dieptes van 40 tot 90 m. De jonge larvale vormen zijn
planktonisch en verplaatsen zich naar water met een lager zoutgehalte om zich als juvenielen te ontwikkelen. Oudere garnalen keren terug naar dieper water
offshore.
Kreeften kennen een directe ontwikkeling: er is geen larvale vorm. Uit het ei komt een piepklein juveniel met dezelfde vorm als het volwassen dier en een compleet stel aanhangsels en somieten. Bij de meeste kreeftachtigen verloopt de ontwikkeling echter indirect en komt uit het ei een larve die qua structuur en uiterlijk sterk verschilt van de volwassene. De overgang van larve naar volwassen dier is de metamorfose. De primitieve en meest voorkomende larve bij de Crustacea is de nauplius (Figuur 19-9 en 19-23). Nauplii dragen slechts drie paar aanhangsels: unirameeuze eerste antennes, birameeuze antennes, en birameeuze onderkaken. Alle fungeren in dit stadium als zwemmende aanhangsels.
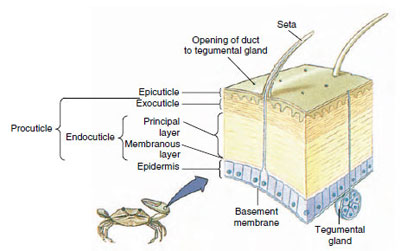
Figuur 19-10
Structuur van schaaldier cuticula.
De verdere ontwikkeling kan een geleidelijke overgang naar de volwassen lichaamsvorm inhouden, waarbij via een reeks vervellingen aanhangsels en somieten worden toegevoegd, of de aanname van de volwassen vorm kan gepaard gaan met meer abrupte veranderingen. De metamorfose van een zeepok bijvoorbeeld gaat van een vrij zwemmende nauplius over in een larve met een tweekleppig kopborststuk, cyprid genaamd, en tenslotte in een sessiele volwassene met kalkplaten.
Ecdysis: Ecdysis (ek´duh-sis) (Gr. ekdyein, afstropen), of vervelling, is nodig om het lichaam in omvang te laten toenemen, omdat het exoskelet niet-levend is en niet groeit naarmate het dier groeit. Veel van het functioneren van een schaaldier, met inbegrip van de voortplanting, het gedrag en veel stofwisselingsprocessen, wordt rechtstreeks beïnvloed door de fysiologie van de vervellingscyclus.
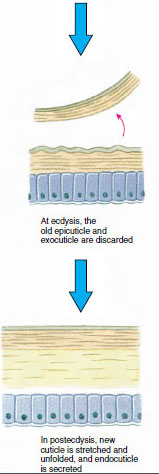
De cuticula, die wordt afgescheiden door de onderliggende opperhuid, bestaat uit verschillende lagen (Figuur 19-10). De buitenste laag is epicuticula, een zeer dun laagje met lipiden geïmpregneerd eiwit. Het grootste deel van de cuticula wordt gevormd door de verschillende lagen procuticula: (1) exocuticula, die vlak onder de epicuticula ligt en eiwit, calciumzouten en chitine bevat; (2) endocuticula, die zelf bestaat uit (3) een hoofdlaag, die meer chitine en minder eiwit bevat en sterk verkalkt is, en (4) een ongekalkte membraneuze laag, een betrekkelijk dun laagje chitine en eiwit.
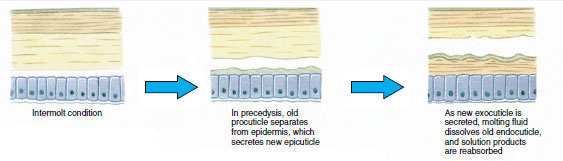
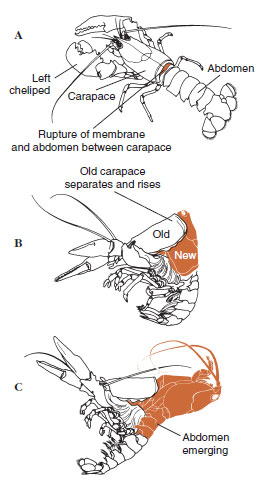
Enige tijd voor de eigenlijke ecdysis, vergroten de epidermale cellen aanzienlijk. Ze scheiden zich af van de membraneuze laag, scheiden een nieuw epicuticulum af en beginnen een nieuw exocuticulum af te scheiden (Figuur 19-11). Enzymen komen vrij in het gebied boven de nieuwe epicuticula. Deze enzymen beginnen de oude endocuticle op te lossen, en de oplosbare producten worden geresorbeerd en opgeslagen in het lichaam van het schaaldier. Sommige calciumzouten worden als gastrolieten (minerale afzettingen) in de wanden van de maag opgeslagen. Uiteindelijk blijven alleen exocuticula en epicuticula van de oude cuticula over, die worden bedekt met nieuwe epicuticula en nieuwe exocuticula. Het dier slikt water in, dat het via zijn darm opneemt, en zijn bloedvolume neemt sterk toe. De inwendige druk doet de cuticula splijten, en het dier trekt zich uit zijn oude exoskelet (Figuur 19-12). Dan volgt een uitrekking van de nog zachte nieuwe cuticula, de afzetting van de nieuwe endocuticula, de herpositionering van de geredde anorganische zouten en andere bestanddelen, en de verharding van de nieuwe cuticula. Tijdens de vervellingsperiode is het dier weerloos en blijft het verborgen.
 |
|
 |
 |
| Figuur 19-12 Molting sequence in a lobster, Homarus americanus. A: Het membraan tussen het kopborststuk en het achterlijf scheurt en het kopborststuk begint langzaam omhoog te komen. Deze stap kan tot 2 uur duren. B en C: Hoofd, borststuk en tenslotte achterlijf trekken zich terug. Dit proces duurt gewoonlijk niet langer dan 15 minuten. Onmiddellijk na de ecdysis zijn de wangslippen uitgedroogd en is het lichaam zeer zacht. De kreeft blijft snel water opnemen, zodat het lichaam binnen 12 uren ongeveer 20% in lengte en 50% in gewicht toeneemt. Het weefselwater zal in de daaropvolgende weken worden vervangen door eiwitten. |
Figuur 19-11 Scheiding en resorptie tijdens ecdysis. |
Wanneer een schaaldier jong is, moet ecdysis vaak plaatsvinden om groei mogelijk te maken, en de vervellingscyclus is betrekkelijk kort. Naarmate het dier de volwassenheid nadert, worden de perioden tussen de vervellingen steeds langer, en bij sommige soorten houdt het vervellen helemaal op. Tijdens de perioden van vervelling neemt de weefselmassa toe doordat levend weefsel het water vervangt.
Hormonale controle van de ecdysiscyclus
Hoewel ecdysis hormonaal gecontroleerd wordt, wordt de cyclus vaak op gang gebracht door een omgevingsprikkel die door het centrale zenuwstelsel wordt waargenomen. Dergelijke prikkels kunnen temperatuur, daglengte en vochtigheid (in het geval van landkrabben) zijn. Het signaal van het centrale zenuwstelsel vermindert de productie van een vervellingsremmend hormoon door het X-orgaan. Het X-orgaan is een groep neurosecretorische cellen in de medulla terminalis van de hersenen. Bij rivierkreeften en andere decapoden bevindt de medulla terminalis zich in de oogsteel. Het hormoon wordt in de axonen van het X-orgaan overgebracht naar de sinusklier (die zelf waarschijnlijk niet kliervormig is), eveneens in de oogsteel, waar het wordt afgegeven aan de hemolymfe.
Een daling van het niveau van het vervellingshormoon bevordert het vrijkomen van een vervellingshormoon uit de Y-organen. De Y-organs bevinden zich onder de opperhuid in de buurt van de adductorspieren van de onderkaken, en zijn homologisch met de prothoracale klieren van insecten, die het hormoon ecdysone produceren. De werking van het vervellingshormoon is het op gang brengen van processen die leiden tot ecdysis (proecdysis). Eenmaal begonnen, verloopt de cyclus automatisch zonder verdere actie van hormonen uit het X- of het Y-orgaan.
Andere endocriene functies: Niet alleen versnelt het verwijderen van de oogstengels het vervellen, meer dan 100 jaar geleden werd ook ontdekt dat kreeftachtigen waarvan de oogstengels zijn verwijderd, hun lichaamskleur niet meer aan de omstandigheden op de achtergrond kunnen aanpassen. Meer dan 50 jaar geleden werd ontdekt dat dit gebrek niet werd veroorzaakt door verlies van gezichtsvermogen, maar door verlies van hormonen in de oogstengels. De lichaamskleur van schaaldieren is grotendeels het resultaat van pigmenten in speciale vertakte cellen (chromatoforen) in de opperhuid.
Concentratie van pigmentkorrels in het centrum van de cellen veroorzaakt een lichter effect, en verspreiding van pigment over de cellen veroorzaakt een donkerder effect. Het pigmentgedrag wordt gestuurd door hormonen uit neurosecretorische cellen in de oogsteel, evenals de migratie van retinaal pigment voor licht- en donkeradaptatie in de ogen (figuur 19-8).
Vrijkomen van neurosecretorisch materiaal uit de pericardiale organen in de wand van het pericard veroorzaakt een toename van de snelheid en amplitude van de hartslag.
Androgene klieren, voor het eerst gevonden bij een amfipode (Orchestia, een gewone strandhopper), komen voor bij mannelijke malacostracans. In tegenstelling tot de meeste andere endocriene organen bij schaaldieren, zijn dit geen neurosecretorische organen. Hun secretie stimuleert de expressie van mannelijke geslachtskenmerken. Jonge malacostracans hebben rudimentaire androgene klieren, maar bij wijfjes komen deze klieren niet tot ontwikkeling. Als ze bij een vrouwtje kunstmatig worden geïmplanteerd, veranderen haar eierstokken in teelballen en beginnen ze sperma te produceren, en haar aanhangsels beginnen bij de volgende vervelling mannelijke kenmerken aan te nemen. Bij isopoda bevinden de androgene klieren zich in de teelballen; bij alle andere malacostracans bevinden zij zich tussen de spieren van de coxopoden van de laatste thoracale poten en zijn zij gedeeltelijk bevestigd bij de uiteinden van de vasa deferentia. Hoewel de vrouwtjes geen organen bezitten die lijken op androgene klieren, produceren hun eierstokken een of twee hormonen die de secundaire geslachtskenmerken beïnvloeden.
Hormonen die andere lichaamsprocessen bij Crustacea beïnvloeden, kunnen aanwezig zijn, en er zijn aanwijzingen dat een neurosecretorische stof die in de oogsteel wordt geproduceerd, de bloedsuikerspiegel regelt.
Voedingsgewoonten
Voedingsgewoonten en aanpassingen aan het voeden variëren sterk onder kreeftachtigen. Veel vormen kunnen van het ene type voeding overschakelen op het andere, afhankelijk van de omgeving en de beschikbaarheid van voedsel, maar ze gebruiken allemaal dezelfde fundamentele set monddelen. Onderkaken en bovenkaken dienen om voedsel op te nemen; bovenkaakpoten houden voedsel vast en pletten het. Bij roofdieren dienen de poten, met name de jukbeenderen, om voedsel te vangen.
Veel grote en kleine kreeftachtigen zijn roofdieren, en sommige hebben interessante aanpassingen om prooien te doden. Een garnaalachtige vorm, Lygiosquilla, heeft aan een van zijn looppoten een gespecialiseerd tandbeen dat in een groef kan worden getrokken en plotseling kan worden losgelaten om een passerende prooi te doorboren. Pistoolgarnalen (Alpheus spp.) hebben een enorm vergrote chela die als de haan van een geweer kan worden gespannen en met een zodanige kracht kan worden afgeklapt dat hun prooi erdoor wordt verdoofd.
Het voedsel van zweefgarnalen varieert van plankton en detritus tot bacteriën. Roofdieren eten larven, wormen, schaaldieren, slakken en vissen. Aaseters eten dood dierlijk en plantaardig materiaal. Zweefgarnalen, zoals de elfengarnaal, watervlooien en zeepokken, gebruiken hun poten, die voorzien zijn van een dikke rand setae (borstelharen), om waterstromen op gang te brengen die voedseldeeltjes door de setae (borstelharen) vegen. Moddergarnalen (Upogebia spp.) gebruiken lange setae op hun eerste twee paar thoracale aanhangsels om voedselmateriaal te zeven uit water dat door hun hol wordt gecirculeerd door bewegingen van hun zwemmersets.
Kreeften hebben een tweedelige maag (Figuur 19-13). Het eerste deel bevat een maagmolen waarin voedsel, dat reeds door de onderkaken is verscheurd, door drie kalkachtige tanden verder kan worden vermalen tot deeltjes die fijn genoeg zijn om door een setosefilter in het tweede deel te passeren; de voedseldeeltjes gaan vervolgens naar de darm voor chemische vertering.
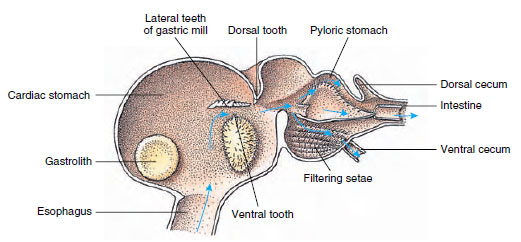
Figuur 19-13
Malacostracaanse maag toont maag-“molen” en richtingen van voedselbewegingen. De molen heeft chitineachtige richels, of
tanden, voor het kauwen, en setae voor het uitrekken van voedsel voordat het in de pylorische maag komt.
Geef een antwoord