甲殻綱
甲殻類の一般性
甲殻類は他の節足動物といろいろ違うが、本当に区別できる特徴は、節足動物で唯一触角が2対あることである。 甲殻類は、触角と大あごに加え、頭部に2対の上顎があり、さらに各体節に1対の付属肢がある。 甲殻類の中には、すべての体節に付属肢がないものもある。 第一触角を除くすべての付属肢は原始的に二股であり、現在の成体でも少なくともいくつかの付属肢はその状態である。 呼吸に特化した器官がある場合は、エラとして機能します。
ほとんどの甲殻類は16〜20節ですが、60節以上のものもあります。 体節数が多いのは原始的な特徴である。 より派生的には、体節数が少なく、タグマティシティが大きくなる。 主なタグマタは頭部,胸部,腹部であるが,頭胸部のように体節の融合の程度が異なるため,クラス全体(あるいはいくつかの亜クラス内)で相同ではない。
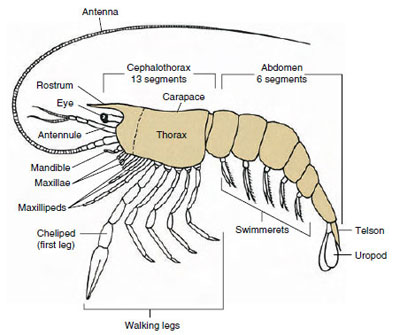
Figure 19-1
Archetypical plan of Malacostraca. 710>
甲殻類の中で圧倒的に多いのがマラコストラカ綱で、ロブスター、カニ、エビ、ビーチホッパー、ダンゴムシなど多くの種類が含まれます。 これらは驚くほど一定の体節とタグマタの配置を示し、このクラスの祖先のプランと考えられている(図19-1)。 この典型的な体型は、5体節(胚発生時には6体節)の融合した頭部、8体節の胸部、6体節(数種では7体節)の腹部からなる。 前端には非分節の吻があり、後端には非分節の尾節があり、腹部最終体節とそのウロポッドとともに尾扇を形成するものが多い。
多くの甲殻類では、頭部の背面甲殻が後方および側方に伸びて胸部および腹部の一部または全部を覆うか融合していることがある。 このような被覆を甲羅という。 甲羅は蛤の殻のような弁を形成し、体の大部分または全部を覆っているグループもある。 十脚類(ロブスター、エビ、カニなど)では、甲羅は頭胸部全体を覆うが、腹部は覆わない。
形態と機能
ザリガニなどの大型甲殻類は、その大きさと入手しやすさから、他のグループより多く研究されてきた。 また、実験入門コースでもよく研究されている。
外見的特徴
甲殻類の体は、キチン質、タンパク質、石灰質からなる分泌性のクチクラで覆われている。 特に大型の甲殻類の硬くて重い板には、石灰質が多く含まれています。 硬い保護膜は、体節間の接合部では柔らかく薄いので、柔軟な動きが可能である。 甲羅がある場合は頭胸部の大部分またはすべてを覆う。ザリガニなどの十脚類では、頭部と胸部のすべての節が背面から甲羅に包まれている。 甲羅に囲まれていない各体節は背面のクチクラ板で覆われ(図19-2A)、腹部の横棒である胸骨が各体節付属物の間にある(図19-2B)。 腹部は距骨で終わるが、距骨は体節とはみなされず、肛門を持つ。 (距骨は環形動物の錐体と相同なのかもしれない)
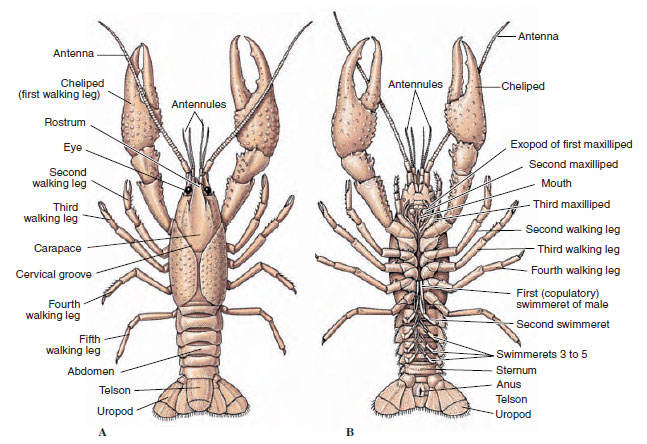
図19-2
ザリガニの外部構造。 A、背面図。 B、腹面図。
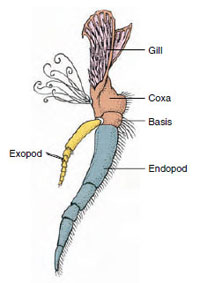
図19-3
甲殻類の二枚貝
付属器(
crayfishの第3上顎骨)
性とグループにより鰓孔の位置は異なる。 性差やグループによって生殖孔の位置は異なり、一対の付属器上や基部にあったり、体の末端にあったり、脚のない体節上にあったりする。 ザリガニの場合,精管は第5歩脚の基部中央,卵管は第3歩脚の基部に開口する。 雌では精嚢の開口部は通常、第4対の歩行脚と第5対の間の腹部中央線に位置する。 ザリガニ綱とレミペディア綱の魚類は、通常、各体節に一対の関節をもつ付属肢をもつが(図19-3)、他の綱の腹部体節には付属肢をもたない。 ザリガニのような派生した甲殻類の付属器には、かなり特殊なものがあることがわかる。 しかし、いずれも基本的な二層式プランの変形であり、ザリガニの付属器であるmaxilliped(胸部の肢が頭部付属器となったもの)などで説明できる(図19-3、図19-4)。 基部である原脚には、側方の外脚と内側の内脚がある。 原脚は1〜2個の関節(基節と尾節)からなり、外脚と内脚はそれぞれ1〜数個の関節をもつ。 ザリガニの歩行脚のように、二次的に単弓型になる付属器もある。 甲殻類の肢には内方突起や側方突起が生じることがあり、それぞれ内脚、外脚と呼ばれるが、原脚の外脚は上脚と呼ばれる。 上脚はしばしば鰓(えら)として変化する。 表19-1は、様々な付属物が、特定の機能に適合するように二足歩行プランから変更されるようになったことを示す。
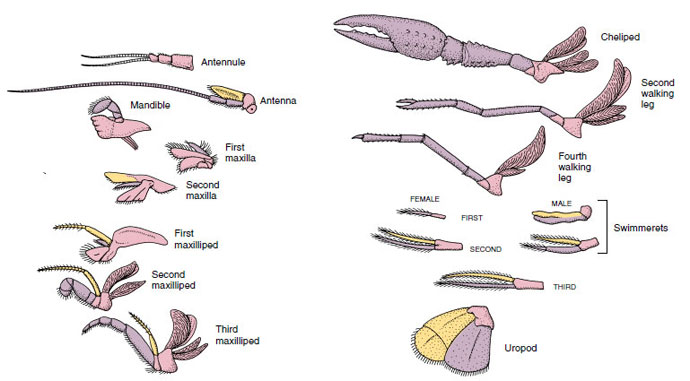
図19-4
ザリガニの付属器で、
スイムメレットのように基本2角形のプランからどのように変化してきたかを示したものです。 前脚はピンク、内脚は紫、外脚は黄色。
基本プランが似ていて、共通の形態から派生した構造は、機能が同じかどうかにかかわらず、相同であると言われます。 特殊な歩行脚、口器、鉤脚、遊泳脚はすべて共通の二又型から発達しましたが、異なる機能を果たすように変化したので、それらはすべて互いに相同であり、直列相同性として知られている状態です。 原始的に四肢はすべて非常によく似ていたが、構造的な改変の進化の過程で、ある枝は縮小され、あるものは失われ、あるものは大きく変化し、あるものは新しい部分が追加されたのである。 ザリガニとその仲間は動物界で最も精巧な系列相同性を持ち、17種類の異なるが系列的に相同な付属器を持っている(表19-1)
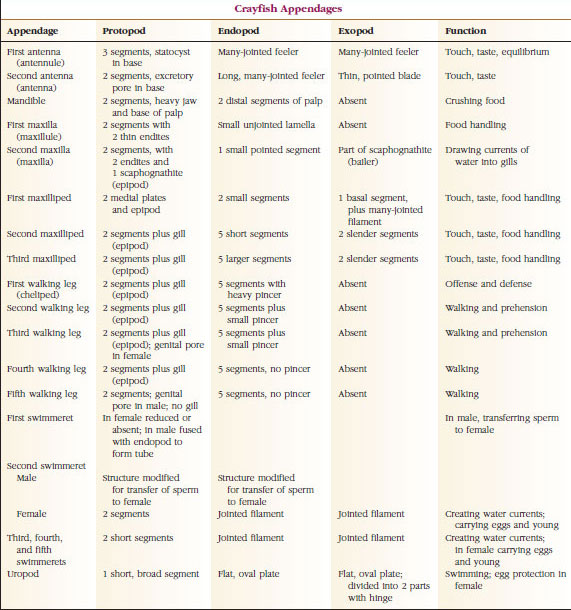
TABLE 19.1
内部の特徴
筋肉系や神経系、胸部や腹部の分節は明らかに環形動物の祖先から受け継いだメタメリズムを示しているが、その他の系には著しい変化がある。
血行:節足動物の主要な体腔は腔ではなく、血液で満たされた血行管である。 ほとんどの節足動物の胚発生期には,少なくともいくつかの体節の中胚葉の中に胎生期の腔が開口している。 これらはすぐに消滅するか、発達中の中胚葉および外胚葉の構造と卵黄の間の空間と連続するようになる。 この空間は血流管となり、中胚葉の腹膜で覆われることはない。 甲殻類では、骨格区画は排泄器官の末端嚢と生殖腺の周りの空間しか残っていない。 筋系:甲殻類の多くは筋が体のかなりの部分を占めている。 筋肉は通常、部位を体側に引き寄せる屈筋と、外側に伸ばす伸筋という、拮抗するグループに分かれている。 ザリガニの腹部には強力な屈筋があり(図19-5)、これはザリガニが後方に泳ぐとき、つまり最高の逃走手段に使われる。 胃の両側にある強い筋肉は大あごを制御する。
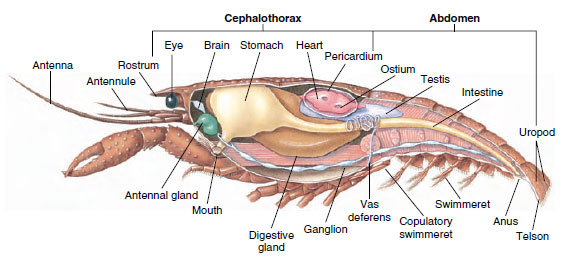
図19-5
オスザリガニの内部構造
呼吸器系. 小型の甲殻類の呼吸ガス交換は、クチクラの薄い部分(例えば、脚部)または体全体で行われ、特殊な構造は存在しないこともある。 大型の甲殻類にはエラがあり、これは非常に薄いキューティクルを持つ繊細な羽毛状の突起である。 十腕類では甲羅の側面が鰓孔を囲み、前方および腹側に開口している(図19-6)。 鰓は胸壁から鰓腔内に突出するもの、胸脚と胴体の連結部から突出するもの、胸部胸郭から突出するものがある。 後者2種はザリガニ類に典型的である。 第2上顎骨の一部である “bailer “は、水を鰓孔に引き込み、脚の付け根で鰓孔に、前方で鰓孔から排出する
循環器系。 甲殻類などの節足動物は、「開口型」または「ラクナ型」の循環器系を持っている。 つまり、閉鎖系の動物のように静脈がなく、血液と間質液の分離がない。 血液は動脈を通って心臓を出て、血球を循環し、静脈の代わりに静脈洞(空間)に戻って心臓に再び入る。 背側の心臓は、脊椎動物と同じように閉じたシステムになっています。 背側の心臓は、筋状の筋肉からなる単室性の袋である。 血流は周囲の心膜洞から一対の通路を通って心臓に入り、洞への逆流を防ぐ弁がある(図19-6)。 心臓から血流は一つまたは複数の動脈に入る。 動脈には弁があり、血液リンパの逆流を防いでいる。 小動脈は組織の洞に流れ込み、その洞はしばしば大きな胸骨洞に流れ込む(図19-6)。
そこから求心性の洞チャネルが、酸素と二酸化炭素の交換のために、存在すれば鰓に血リンパを運ぶ。
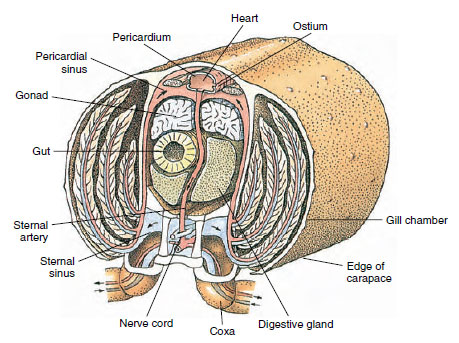
図19-6
この「開いた」
血液システムの血流方向を示すザリガニの心臓部の図示された断面図です。 心臓は動脈を通して体組織に血液を送り、その血液は組織の洞に流れ込みます。 戻った
血液は胸骨洞に入り、ガス交換のためにエラを通り、最後に求心性の
チャネルによって心膜洞に戻る。
節足動物の血流は大部分が無色である。 少なくとも2種類のアメーバ状の細胞が含まれている。 銅を含む呼吸色素であるヘモシアニンや鉄を含む色素であるヘモグロビンが溶液中で運ばれていることがある。 ヘモリンパは凝固する性質があり、軽傷で失われるのを防ぐことができる。
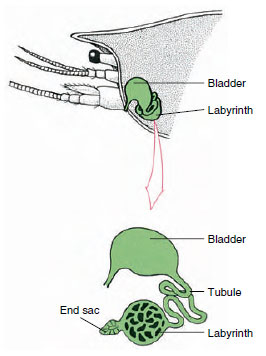
図19-7
ザリガニの触角腺(緑腺)の模式図です。 (甲殻類の中には迷路がなく、
排泄管(ネフリッド管)は
コイル状の管であるものがある。 甲殻類の成体の排泄器官は食道より前方の頭部腹側にある一対の管状の構造である(図19-5)。 触角の基部で開くか、第二上顎の基部で開くかによって、触角腺または上顎腺と呼ばれる。 成体の甲殻類では、両方を持つものも少なくない。 十脚類の排泄器官は触角腺で,この仲間では緑腺とも呼ばれる。 甲殻類はクモや昆虫の排泄器官であるマルピーギ管を持っていない。
胚の骨格区画に由来する触角腺の末端嚢は、小さな小胞(サキュール)と迷路というスポンジ状の塊から構成されている。 迷路は排泄管によって背側膀胱につながり、背側膀胱は触角基底部の腹面にある孔によって外部に開口している(図19-7)。 血流管内の静水圧は、液体を末端嚢に濾過する力を提供する。 濾液は排泄管と膀胱を通過する際に、塩類、アミノ酸、グルコースおよび若干の水分が再吸収され、最終的に尿として排泄される。
窒素性廃棄物(主にアンモニア)の排出は、特に鰓などの薄い角質層を拡散して行われ、いわゆる排泄器官は主に体液のイオンと浸透圧組成を調節する機能を有する。 ザリガニのような淡水性の甲殻類は、エラなどの水透過性表面を伝って拡散する水による血液の過剰希釈に常に脅かされている。 緑色腺は、塩分の少ない希薄な尿を形成することで、効果的な「洪水防止装置」として機能する。 尿中にNa+とCl-が失われるが、この損失はエラによる溶存塩の積極的な吸収によって補われる。 ロブスターやカニなどの海産甲殻類では、腎臓は尿細管の塩分を選択的に変化させることによって、血流の塩分組成を調整する機能を持つ。 これらの形態では、尿は血液と等浸透圧のままである。
神経・感覚系 甲殻類と環形動物の神経系は共通点が多いが,甲殻類では神経節がより融合している(図19-5)。 眼と2対の触角に神経を供給する一対の食道上神経節である。 口、付属器、食道、触角腺に神経を供給する少なくとも5対の神経節が融合した食道下神経節と連結部で結ばれている。
甲殻類は環形動物に比べ感覚器が発達しており、体節ごとに1対の神経節があり、付属器や筋肉などに神経を供給しています。 甲殻類の最大の感覚器は眼と静止胞です。 体全体に広く分布する触毛は、キューティクルの繊細な突起で、特に蹄鉄、口器、距骨に多く見られます。 味覚や嗅覚などの化学感覚は触角や口器などの毛にあります。
ザリガニの第一触角の基節には、背中の孔で表面に開いている袋状の静止嚢があります。 静止胞の中には、キチン質でできた感覚毛と静止石となる砂粒が付着した隆起部がある。 動物が姿勢を変えるたびに、感覚毛の砂粒の位置が変化し、それが刺激として脳に伝わり、動物はそれに合わせて姿勢を変えることができる。 クチクラの脱皮のたびに、静止胞体のクチクラの裏地は失われ、砂粒も一緒に失われる。
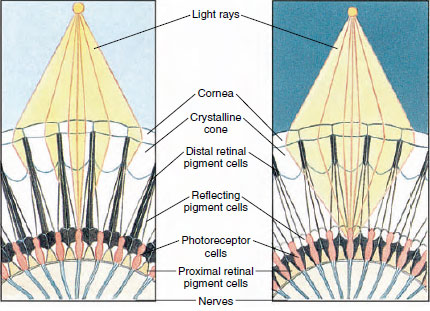
図19-8
節足動物の複眼の一部で、昼と夜を見るために色素
を移動させているところ。 各図には5つの扁桃体が描かれています。 昼間は各オマチダが暗い
色素襟で囲まれているので、各オマチダは自分の角膜に入った光線
だけが刺激される(モザイク視)。夜間は色素が
不完全な襟を作り、光線は隣のオマチダ
に広がる(連続、あるいは重なり合い像)。
多くの甲殻類の目は複眼で、omatidiaという多数の視細胞単位で構成されています(図19-8)。 それぞれの眼の丸い表面を覆っているのは透明なクチクラで、角膜は切子と呼ばれる多くの小さな正方形または六角形に分かれている。 これらの面は、オマティディアの外側の端である。 各オマチダは小さな目のような働きをし、数種類の細胞が柱状に並んでいる(図19-8)。 節足動物の複眼では、色素が移動することによって、さまざまな光量に対応することができる。 各オマティジウムには、遠位網膜、近位網膜、反射の3組の色素細胞があり、これらは各オマティジウムの周りに多かれ少なかれ完全なカラーまたはスリーブを形成できるように配置されている。 強い光や昼間の適応のためには、遠位網膜色素は内側に移動し、外側に移動する近位網膜色素と出会うので、完全な色素スリーブが眼球のまわりに形成される(図19-8)。 この状態では、角膜に直接当たった光線だけが視細胞(網膜細胞)に到達することになる。 そのため、各子葉は視野の限られた範囲しか見えない(モザイク像、または貼付像)。 薄暗いところでは、遠位と近位の色素が分離し、光線は反射する色素細胞の助けを借りて、隣接するオーマット膜に広がり、連続した像(重ね合わせ像)を形成する機会を持つ。
生殖、ライフサイクル、内分泌機能
ほとんどの甲殻類は雌雄が分かれており、グループによって交尾にさまざまな特殊性があります。 フジツボは単為生殖だが、一般に他家受精を行う。 貝類では雄が少なく,単為生殖が一般的であるものもある。 枝角類やフジツボ類は特殊な産卵室を持ち、橈脚類は腹部側面に産卵袋を持ち(図19-19参照)、多くの甲殻類は腹部付属肢に卵や子を付着させている。
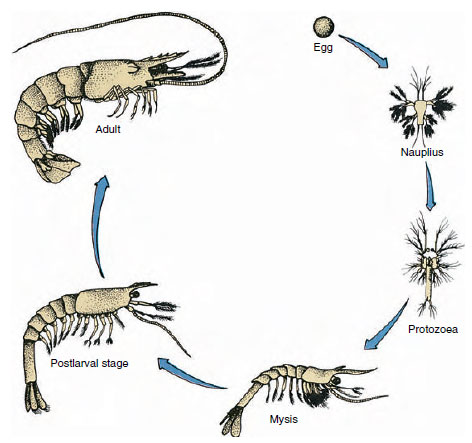
図19-9
ガルフシュリンプPenaeusのライフサイクル。 水深40~90mで産卵した幼生は
浮遊性で、塩分濃度の低い海域に移動して稚魚となる。 710>
カニ類は直接発生で、幼生形態はない。 卵から成魚と同じ形、付属器や体節がそろった小さな稚魚が孵化します。 しかし、多くの甲殻類は間接的な発生であり、卵から成虫とは全く異なる構造・外観の幼生が孵化する。 幼虫から最終的に成虫になるのが変態である。 甲殻類で最も原始的で広く見られる幼虫は、ナウプリウスです(図19-9、図19-23)。 単弓類の第1触角、双弓類の触角、双弓類の大あごの3対の付属器しか持たない。
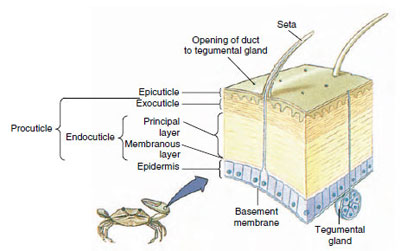
図19-10
甲殻類のキューティクルの構造。
その後の発生は、脱皮を繰り返して付属肢や体節が追加され、徐々に成体へと変化する場合と、成体への移行がより急激な変化を伴う場合がある。 例えば、フジツボは、自由遊泳するナウプリウスから、イプリッドと呼ばれる二枚貝の甲羅を持つ幼生になり、最後に石灰質板を持つ無柄の成体に変態する
。 外骨格は非生物であり、成長とともに大きくならないため、体を大きくするためには脱皮が必要である。 甲殻類の生殖、行動、多くの代謝過程などの機能の多くは、脱皮サイクルの生理に直接影響されます。
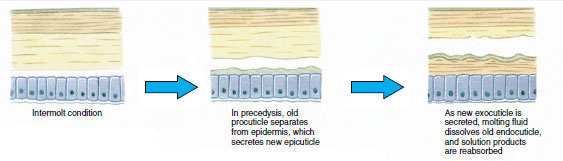
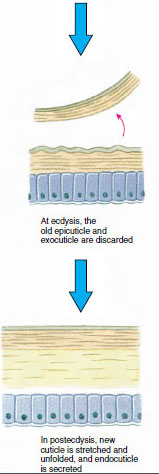
表皮の下に分泌されるキューティクルは、いくつかの層になっています(図19-10)。 最も外側にあるのはエピキューティクルで、脂質を含んだタンパク質の非常に薄い層である。 キューティクルの大部分はプロキューティクルの数層である。 (1)外皮は上皮のすぐ下にあり、タンパク質、カルシウム塩、キチンを含む。(2)内皮はそれ自体、(3)キチン質を多く含み、タンパク質が少なく、石灰化が激しい主層と、(4)キチン質とタンパク質からなる比較的薄い層の未石灰化の膜状層に構成されている。 膜層から分離し、新しいエピキューティクルを分泌し、新しいエキソキューティクルを分泌し始める(図19-11)。 新しいエピキューティクルの上の領域には酵素が放出される。 この酵素は古いエンドキューティクルを分解し始め、可溶性の生成物は甲殻類の体内に再吸収され貯蔵される。 カルシウム塩の一部は胃壁にガストロリス(鉱物の付着物)として貯蔵される。 最後に、古いキューティクルの外皮と上皮だけが残り、新しい上皮と新しい外皮に覆われる。 水を飲み込み、腸から吸収し、血液量が大きく増加する。 内圧でキューティクルが裂け、古い外骨格から身を引き抜く(図19-12)。 その後、まだ柔らかい新皮質の伸展、新内皮の沈着、回収した無機塩類などの成分の再沈着、新皮質の硬化が行われる。 脱皮の期間中は無防備になり、隠れている。
 |
|
 |
 |
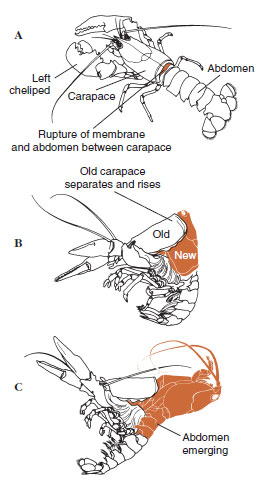
| Figure 19-12 Homarus americanus, lobsterにおける一連の脱皮。 A,甲羅 と腹部の間の膜が破れ、甲羅がゆっくりと上昇し始める。 この段階には最大で2時間かかることもある。 BとC、頭部、胸部、そして最後に腹部が引っ込む。 この工程は通常 15分もかからない。 脱皮直後は足が乾燥し、体も非常に柔らかい。 その後、ロブスターは急速に水を吸収し、12時間以内に体長は約20%、体重は約50%増加する。 |
Figure 19-11 脱皮におけるキューティクルの分泌と 再吸収 |
甲殻類が若いうちは、成長のために頻繁に脱皮しなければならず、脱皮周期は比較的短いです。 成熟期に近づくと脱皮周期は次第に長くなり、脱皮が完全に停止する種もある。
ホルモンによる脱皮の制御
脱皮はホルモンによって制御されているが、中枢神経系が感知する環境刺激によって開始されることが多い。 そのような刺激には、温度、日長、湿度(陸ガニの場合)などがある。 中枢神経系からの信号により、X器官からの脱皮抑制ホルモンの分泌が減少する。 X器官とは、脳の終末髄質にある神経分泌細胞群である。 ザリガニなどの十脚類では、終末髄質が眼柄に存在する。 脱皮ホルモンが低下すると、Y器官から脱皮ホルモンが放出される。 Y器官は大顎内転筋付近の表皮下にあり、昆虫の前胸腺と同質のもので、エクジソンというホルモンを分泌する。 脱皮ホルモンの作用は、脱皮に至るプロセスを開始させることである(前駆脱皮)。 一度脱皮が始まると、X器官やY器官からホルモンが分泌されることなく、自動的に脱皮サイクルが進行します。 眼柄を切除すると脱皮が促進されるだけでなく、眼柄を切除された甲殻類は、背景の状況に応じて体色を調整できなくなることが100年以上前に判明しています。 この現象は、視力の低下ではなく、眼柄のホルモンの減少によるものであることが、50年以上前に判明している。 甲殻類の体色は、表皮にある特殊な分岐した細胞(色素細胞)の色素が大きな役割を果たしており、
色素顆粒が細胞の中心に集中すると明るくなり、全体に分散すると暗くなる。
心膜の壁にある心膜器官から神経分泌物質が放出されると、心拍数と振幅が増加する。
両生類(Orchestia、ビーチホッパー)で最初に見つかったアンドロゲン腺は、マラコストラクスの雄に存在する。 甲殻類の他の多くの内分泌器官とは異なり、これらは神経分泌器官ではありません。 この分泌物が雄の性徴の発現を刺激する。 若いマラコストラクサンは、初歩的なアンドロゲン腺を持っていますが、メスではこれらの腺は発達しません。 この腺を人工的に埋め込むと、雌の卵巣は精巣に変化して精子を生産し始め、次の脱皮で付属器官は雄の特徴を持ち始める。 等脚類では、アンドロゲン腺は精巣にありますが、他のすべての悪脊椎動物では、最後の胸脚の尾脚の筋肉の間にあり、一部は精索静脈の端の近くに付着しています。
甲殻類の他の身体プロセスに影響を与えるホルモンが存在する可能性があり、眼肋部で作られる神経分泌物質が血糖値を調節していることを示す証拠があります。 環境や餌の有無によって、摂食形態を変えるものも多いですが、基本的な口器はすべて同じものを使っています。 大あごや上顎は食物を摂取するために機能し、上顎は食物を保持し、粉砕する。
大小多くの甲殻類が捕食性であり、中には獲物を殺すための興味深い適応を持つものもいる。 エビの仲間であるリギオスキーラは、歩行脚の片方に特殊な指があり、これを溝に引き込み、突然放つことで通過する獲物に突き刺すことができるのだそうです。 ピストルシュリンプ(Alpheus spp.)は、巨大化したシェラを銃のハンマーのように構え、獲物を気絶させる勢いで折ることができる。
懸垂餌類はプランクトンやデトリタス、バクテリアなど幅広い餌を持っている。 捕食者は、幼虫やミミズ、甲殻類、カタツムリ、魚類などを食べます。 スカベンジャーは、動物や植物の死骸を食べます。 フェアリーシュリンプ、ミジンコ、フジツボなどの懸濁食は、脚にある太い節で水流を作り、その流れに乗って餌の粒を掃き出す。 マッドシュリンプ(Upogebia spp.)は、胸部付属器の最初の2対にある長い節足動物を使って、泳毛の動きによって巣穴の中を循環する水から餌の材料を漉す。
クレパス類は、胃が2つに分かれている(図19-13)。 第1部には胃粉砕機があり、大あごで引き裂かれた食物は3本の石灰質の歯でさらに粉砕され、第2部の瀬戸内フィルターを通過できるほど細かい粒子となり、食物の粒子は腸に入り、化学的に消化される。
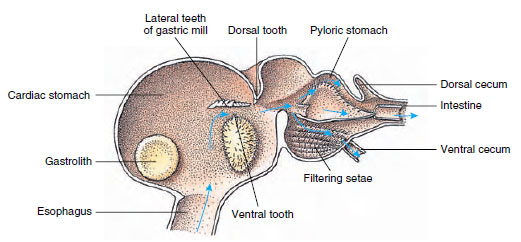
Figure 19-13
Malacostracan stomach showing gastric “mill” and directions of food movement. ミルは咀嚼のためのキチン質の隆起(
歯)と、幽門胃に入る前に食物を絞るための棘突起を持っている。
コメントを残す