Dit hoofdstuk is relevant voor Sectie G4(ii) van de 2017 CICM Primary Syllabus, waarin van de examenkandidaat wordt verwacht dat hij “de verdeling van het bloedvolume en de bloedstroom in de verschillende regionale circulaties beschrijft … inclusief autoregulatie …. Deze omvatten, maar zijn niet beperkt tot, de cerebrale en ruggenmerg-, lever- en splanchnische, coronaire, renale en utero-placentale circulaties”. De levercirculatie is in de afgelopen papers vijf keer aan de orde geweest (vergeleken met vier keer voor de cerebrale circulatie, waardoor deze 25% belangrijker is). Historische SAQ’s zijn onder andere:
- Vraag 13 uit het tweede paper van 2016
- Vraag 3 uit het tweede paper van 2015
- Vraag 18 uit het eerste paper van 2013
- Vraag 11 uit het eerste paper van 2012
- Vraag 4(p.2) uit het eerste artikel van 2008
Samengevat:
- Leverbloedtoevoer:
- Van de leverslagader (een tak van de coeliacale stam)
- Onder aortadruk (MAP ~65-90 mmHg)
- 30-40% van de bloedstroom (SvO2= 95%; 40-50% van de DO2)
- Van de poortader
- Samenvloeiing van mesenteriale en splenische venen
- Ventieloos, lagedruk veneus systeem (8-10 mmHg)
- 70% van de totale bloedstroom (SaO2=85%; 50-60% van de DO2)
- Totale hepatische bloedstroom: 25% van de totale cardiale output, of 1200ml/min.
- Dit is ongeveer 100ml/100g weefsel/minuut
- Hepatisch zuurstofverbruik is 6ml/100g/min
- Hepatisch veneuze zuurstofverzadiging is ~ 65% normaal
- Hepatische microcirculatie:
- bestaat uit de anastomose van hepatische arteriolen en portale venulen
- Deze vaten komen samen om hepatische sinusoïden te vormen
- Sinusoïden zijn sterk gemodificeerde haarvaten van groot kaliber met discontinu endotheel
- Unieke kenmerken:
- Lage druk, om retrograde stroming in het ventielloze poortsysteem te voorkomen
- Lage stroomsnelheid, om extractie van zuurstof en andere interessante moleculen te bevorderen
- Regulering van leverbloedstroom
- Regulering van portaalveneuze stroming:
- Stroomsnelheid wordt voornamelijk bepaald door splanchnische arteriële stroomsnelheid
- Weerstand verandert als reactie op:
- Humorale signalen (bijv. catecholamines), in shock
- Lokale endocriene signalen (bv. VIP), die vasodilatatie veroorzaken na een maaltijd
- Hepatic arteriële debietregulatie:
- Standaard arteriële regulatiemechanismen: myogeen, flow(shear)-gemedieerd, geleide vasomotorische reacties, immunologisch gemedieerd door ontstekingsmoleculen.
- Hepatische arteriële bufferrespons: hepatische arteriële flow neemt toe als portaal veneuze flow afneemt, en vice versa.
- Externe factoren die de hepatische bloedstroom beïnvloeden:
- Venous return: beïnvloedt de hepatische veneuze drainage (bv. tijdens positieve druk beademing of hartfalen)
- Cardiac output: beïnvloedt de hepatische arteriële stroom rechtstreeks, en de portale stroom onrechtstreeks (bv. bij hartfalen)
- Shock states and exercise: decrease splanchnic blood flow, both portal and hepatic
Abshagen et al (2015) would make an excellent single reference point to somebody trying to revise this topic, if only it were not paywalled by Springer. Voor de freegan bieden Eipel et al. (2010) in wezen hetzelfde materiaal, zonder kosten. En zoals gewoonlijk, in een gespecialiseerd veld, duikt er een auteur op die verantwoordelijk lijkt te zijn voor het grootste deel van de literatuur, die in dit geval W. Wayne Lautt is van de Universiteit van Manitoba; vrijwel alles van zijn team lijkt goud waard te zijn.
- Arteriële bloedtoevoer naar de lever
- Portaalveneuze bloedtoevoer naar de lever
- Veneuze drainage van de lever
- Hepatische microcirculatie
- De lever als bloedreservoir
- Regulatie van de portaalveneuze bloedstroom
- Regulation of hepatic arterial blood flow
- Variabele zuurstofonttrekking door de lever
- Extrinsieke factoren die de perfusie van de lever beïnvloeden
- Veranderingen in het medicijnmetabolisme als gevolg van veranderingen in de leverdoorbloeding
Arteriële bloedtoevoer naar de lever
De arteriële toevoer naar de lever wordt verzorgd door de eigenlijke leverslagader, een aftakking van de gemeenschappelijke leverslagader (een korte uitloper van de coeliacale stam waaruit ook de gastroduodenale en rechter maagslagaders ontspringen). Zonder de gebruikelijke klacht over de zinloosheid van het tonen van ware anatomische relaties aan de persoon die ze nooit zal zien, presenteert de auteur dit mooie inktvisachtige diagram van Chamberlain (2012):
.jpg)
In de handboeken wordt gezegd dat deze slagader ongeveer 350ml/min zuurstofrijk bloed naar de lever voert, bij een juiste arteriële druk met een MAP van ongeveer 65-90 mmHg. Bij een saturatie van 100% en een standaard anemische IC-patiënt-achtige hemoglobine van 100g/L bedraagt de hepatische arteriële DO2 dus ongeveer 48ml/min. Wegens zijn zelfregulerend vermogen zal het werkelijke debiet in een gegeven slagader echter heel verschillend zijn. Als voorbeeld volgt hier een tabel van Tygstrup et al (1962). De auteurs maten deze waarden rechtstreeks aan gekanuleerde hepatische vaten van menselijke proefpersonen. De gemiddelde waarde was ongeveer 550ml/min, of 35% van de totale leverbloedtoevoer, maar dit was binnen een enorm bereik (van 166ml tot meer dan 1L/min):

Portaalveneuze bloedtoevoer naar de lever
De portaalveneuze circulatie is in wezen een passief stromend riool van nogal giftig zuurstofarm bloed, dat bestaat uit een systeem van kleploze vaten met weinig gladde spieren in hun wanden. De superieure mesenteriale vene en de miltaire vene komen samen achter het pancreaslichaam en vormen de poortader, een kort vetvat met een betrekkelijk onopvallende microstructuur van de wand. De vernielde anatomische kunst hier is gestolen van anatomycorner.com:

Zoals gezegd, is dit een systeem van veneuze vaten die geen kleppen hebben. Waarom, is een vraag die open staat voor discussie. Het is in ieder geval niet iets waar je klinisch onderzoek van hoge kwaliteit voor kunt verwachten, dus het enige wat we hebben is de speculatie van deskundigen. Sommigen wijzen op de noodzaak om een onbelemmerde doorstroming te handhaven met een relatief lage drukgradiënt; zogezegd zouden veneuze kleppen het lumen belemmeren en als weerstand fungeren, wat contraproductief zou zijn. Anderen suggereren dat het geen kleppen nodig heeft, omdat het leeft in de lage-druk omgeving van de buik. In de kuiten bijvoorbeeld verandert de druk in het compartiment voortdurend, waardoor de aderen worden samengedrukt – als er geen kleppen zouden zijn, zou dit retrograde stroming naar de voet veroorzaken, hetgeen volledig contraproduktief zou zijn. In de buik is de compartimentsdruk constant (en normaal gesproken laag), wat betekent dat de poortader kan verwachten dat er een betrouwbare eenrichtingsstroom in stand wordt gehouden zonder kleppen.
De stroom door de portale circulatie wordt voornamelijk aangedreven door de overgebrachte druk van het bloed dat door de splanchnische arteriolen omhoog wordt gestuwd. Dientengevolge is deze stroming niet pulserend, en onder weinig druk. Balfour et al. (1954) maten rechtstreeks portale veneuze drukken van ongeveer 8-10 mmHg bij gezonde(re) patiënten. De vasculaire weerstand hier veroorzaakt een drukdaling van 8-10 mmHg in de poortader, tot 2-4 mHg in de centrale venen, die Lautt e.a. (1967) lokaliseerden in kleine post-sinusoïdale venulen (vaten voorbij de sinusoïdale anastomose die afwateren in de leverader en die ongeveer 2 mm in diameter zijn).

Dus ondanks de lage stuwdruk is dit systeem, omdat de vaatweerstand zeer laag is, in staat enorme bloedstromen te geleiden. De meeste leerboeken zullen iets tussen 800 en 1200 ml/min noemen, en dit zal uiteraard afhangen van aan wiens lever je het vraagt. Brown et al. (1989) vroegen het aan vijfenveertig normale Kaukasische levers en kwamen tot een gemiddelde waarde van 864 ml/min in rugligging, die daalde tot 662 ml/min wanneer de proefpersonen rechtop stonden.
De zuurstofverzadiging van portaal veneus bloed is slechts ongeveer 85%, die nog lager daalt na een maaltijd. Hardin et al. (1963) canuleerden rechtstreeks de poortaders van verdoofde honden en maten een gemiddelde waarde van 81%, hoewel de waarden bij sommigen zo laag als 65% waren. Na een maaltijd daalde deze waarde tot 69-76%. Omdat de bloedstroom door dit systeem zo groot is, blijft de totale flux van zuurstoftoevoer echter hoog. Met behulp van de conventionele vergelijkingen kan men berekenen dat een flow van 800ml/min bij een saturatie van 80% en een Hb van 100 een DO2 geeft van 88ml/min. Dit is ongeveer het dubbele van wat door de leverslagader wordt geleverd. Met andere woorden, de leverslagader draagt slechts voor ongeveer 30-40% bij tot de totale zuurstofvoorziening van de lever, ook al wordt in veel handboeken vermeld dat het een 50:50 verdeling is met de poortader (bv. Dancygier, 2010). De oorsprong van deze 50% waarde is waarschijnlijk het oude artikel van Tygstrup et al (1962). Zij rapporteerden talrijke interessante metingen bij de mens (bv. gemiddelde drukken in de levervaten, hun bloedstromen, hun weerstand, enz.), en dit heeft hun artikel tot een aantrekkelijke referentie gemaakt voor verschillende generaties auteurs van handboeken.
Veneuze drainage van de lever
Uit deze vreemde dubbele bloedtoevoer ontvangt de lever een enorme totale bloedstroom van ongeveer 1200-1800 ml/min, wat uiteindelijk neerkomt op ongeveer 20-25% van de cardiale output. Logischerwijze is de uitstroom van leverbloed gelijk aan deze instroom, en de leveraders zijn voldoende groot. Gewoonlijk zijn er drie (rechts, midden en links), maar de anatomie van deze aders blijkt van persoon tot persoon aanzienlijk te variëren, hetgeen een probleem vormt voor anatomen die hebben besloten deze aders te gebruiken om de segmenten van de lever af te bakenen. Overal waar de anatomische rangschikking fabrieksnorm is, is de rechter leverader gewoonlijk dominant, en neemt deze het grootste deel van de veneuze drainage voor zijn rekening.
De lever onttrekt ongeveer 6ml/100g/min zuurstof aan zijn dubbele bloedtoevoer, die gemiddeld 16ml/100g/min zuurstof levert (Lutz et al, 1975). Dit geeft een zuurstofonttrekkingsratio van ongeveer 37%. Hieruit zou je een leverveneuze zuurstofverzadiging verwachten van iets van 60%, wat bijna precies is wat werd gemeten door Finnnerty et al (2019). Of althans, dat is het soort getal dat je zou kunnen verwachten als alles goed is. Zoals hieronder zal worden uitgelegd, varieert de zuurstofonttrekking aanzienlijk, afhankelijk van de toereikendheid van het aanbod en de grootte van de vraag.
Hepatische microcirculatie
Dit verdient hier een vermelding omdat het vrij uniek is vanuit een circulatoir perspectief. Het zou verleidelijk zijn om hier diep in dit onderwerp te duiken, maar voorlopig wordt de lezer in plaats daarvan doorverwezen naar uitstekende gratis artikelen als Wake & Kato (2015). In het kort, portale venules en lever arteriolen fuseren anastomotisch in hepatische sinusoïden, die vervolgens draineren in post-sinusoïdale venules.
De eindvaten van het portale veneuze netwerk behouden een lage weerstand, zelfs tot op een zeer smal kaliber, hetgeen betekent dat het grootste deel van de druk van de portale vene rechtstreeks wordt doorgegeven aan de hepatische sinusoïden. Deze sinusoïden zouden in elk ander orgaan “capillairen” kunnen worden genoemd, maar zij hebben een heel andere structuur: hun diameter is veel groter dan die van een normaal capillair, en zij hebben een discontinu epitheel. De drukgradiënt over deze vaten is betrekkelijk laag; volgens Henriksen & Lassen (1988) is deze onder normale omstandigheden niet meer dan 3-5mmHg. Met zo’n lage stuwdruk heeft de stroming hier een ongewoon lage snelheid, waardoor maximale extractie van zuurstof en andere moleculen mogelijk is. De lage druk helpt ook om de drukgradiënt tussen de portale circulatie en de sinusoïden in stand te houden, waardoor dit kleploze systeem tegen retrograde stroming wordt beschermd.
De lever als bloedreservoir
In de leerboeken wordt veel aandacht besteed aan de opslagfunctie van de lever. Het is een zwaar met bloed gevuld orgaan, dat voor ongeveer 25% uit bloed bestaat (Greenway & Stark, 1971), zoals te zien is in deze afgietsel van het poortsysteem (Okudaira, 1991), het zwart-wit origineel gekleurd in schreeuwerig rood, vermoedelijk voor een soort bloedachtig effect:

Als men een bloedvatenstelsel zou ontwerpen voor een zo actief en ongevalgevoelig organisme als de mens, zou men in de verleiding kunnen komen om dit enorme reservoir van bloed toegankelijk te maken voor het lichaam in tijden van bloeding of inspanning. Dat is in feite wat bij veel zoogdieren gebeurt. Bij de hond bijvoorbeeld kon Guntheroth & Mullins (1963) de mobilisatie aantonen van een opgeslagen hepatosplenisch volume dat gelijk is aan 8% van de totale circulatie, wanneer catecholamine vrijkomt. Andere dierstudies komen in het algemeen tot soortgelijke bevindingen, en hoewel er geen gegevens bij de mens lijken te zijn die dit ondersteunen, zijn de leerboeken het er in het algemeen over eens dat dit waarschijnlijk ook bij de mens gebeurt, en beschrijven de lever als een belangrijk opslagorgaan.
Regulatie van de portaalveneuze bloedstroom
Vanuit de besprekingen hierboven zou men in de val kunnen lopen om te denken dat de portaalveneuze weinig rol speelt bij het regelen van de eigen stroom. In dat geval zou het natuurlijk facetachtig zijn om dit hoofdstuk de titel “regulering van de poortaderlijke bloedstroom” te geven, indien deze volledig ongereguleerd zou zijn. De poortader zou dus worden beschouwd als een dom orgaan dat fungeert als een passief kanaal voor bloed, dat niet in staat is iets intelligenter te doen dan een stolsel te vormen om zichzelf te blokkeren. Dat is natuurlijk niet het geval.
Het is waar dat de doorstroming in de poortader hoofdzakelijk wordt bepaald door de doorstroming in de splanchnische slagaders, die de hoeveelheid bloed bepalen die aan het poortsysteem wordt geleverd. Hieruit volgt logischerwijze dat de portale bloedstroom vatbaar zou moeten zijn voor manipulatie door de vaatweerstand van de splanchnische arteriële circulatie te wijzigen. Dit blijkt inderdaad het geval te zijn, aangezien splanchnische vasoconstrictoren (zoals terlipressine) de portale veneuze doorstroming verminderen. In een studie van Baik et al. (2005) verminderde een dosis terlipressine van 2 mg de stroom in de poortader met bijna 40%, hetgeen de basis is van het therapeutisch effect bij de beheersing van varicesale bloedingen.
De poortader heeft dus wel degelijk gladde spieren en receptoren voor alle belangrijke vasoactieve stoffen. Richardson & Withrington (1981) somt een hele reeks vasopressoren op, en Blei (1989) somt een groot aantal vasodilatoren op, waarvan de volgende lijst een conservatieve verkorting is:
| Vasoconstrictoren | Vasodilatoren |
|
|
Dus reageert de portale veneuze circulatie op een verscheidenheid van stimuli, waarvan sommige de weerstand kunnen verdubbelen of halveren (wat, toegegeven, niet veel zegt, aangezien deze om te beginnen zeer laag is). De reactie op endogene vasopressoren houdt waarschijnlijk verband met de duidelijke rol van de lever als bloedreservoir, in welk geval het zinvol zou zijn het portale veneuze volume te verlagen en het extra bloed naar de systemische circulatie te “spoelen”. De oplettende lezer zal in de bovenstaande lijst ook enkele splanchnische hormonen hebben gezien, die zouden kunnen wijzen op een soort reguleringsmechanisme dat verband houdt met de spijsvertering. Dit is inderdaad het geval. Dauzat et al. (1994) konden dit bij gezonde vrijwilligers onderzoeken met behulp van niet-invasieve meettechnieken, en ontdekten dat de doorsnede van de poortader met 40% toeneemt na een “standaardmaaltijd” (blijkbaar is dat 470 ml Ensure), hetgeen gepaard ging met een enorme toename van 80% van de doorstroming.
Regulation of hepatic arterial blood flow
De leverslagader, een gespierd lid van de systemische circulatie, wordt beïnvloed door allerlei duidelijk omschreven regulerende mechanismen. Als men ze zou moeten classificeren, zouden ze in twee rommelig overlappende categorieën vallen:
- Intrinsieke arteriële autoregulatiemechanismen, die gemeenschappelijk zijn voor alle slagaders in de gehele systemische circulatie
- Hepatische arteriële bufferrespons, die uniek is voor de levercirculatie.
Arteriële autoregulatoire mechanismen worden elders meer in detail besproken, aangezien zij tamelijk generiek zijn en van toepassing zijn op alle arteriële regionale circulatiesystemen. Deze generieke factoren kunnen verder worden ingedeeld in lokale en systemische:
- Systemische factoren omvatten:
- Arteriële baroreflexcontrole (verhoogde bloeddruk leidt tot een daling van de SVR)
- Perifere en centrale chemoreceptoren (hypoxie leidt tot een verhoogde SVR)
- Hormonen (bijv. vasopressine en angiotensine)
- Temperatuur (hypothermie leidt tot verhoogde SVR)
- Lokale/regionale factoren omvatten:
- Intrinsieke myogene regulatie (als reactie op rek)
- Metabole regulatie (als reactie op toegenomen weefselvraag)
- Flow- of shear-geassocieerde regulatie (als reactie op toegenomen lokale flow)
- Geleide vasomotorische reacties van naburige vasculaire sites
- Lokale afkoeling (die eerst tot vasoconstrictie leidt, en dan weer tot vasodilatatie)
- Immunologische modulatie door ontstekingsmediatoren
De leverarteriële bufferrespons is ook bekend onder de welluidende naam, “leverarteriële-portaalveneuze semi-reciprocale interrelatie”. Het basisprincipe kan zeer eenvoudig worden samengevat. Wanneer de portaal veneuze stroom daalt, stijgt de lever arteriële stroom. Met andere woorden, de leverarteriële vaatweerstand is evenredig met de portaalveneuze bloedstroom. Lautt et al. (1990) konden aantonen dat deze relatie relatief lineair is, over een normaal bereik van debieten:
.jpg)
Deze relatie werkt in een vrij snel tijdsbestek. Wanneer de poortader intraoperatief wordt afgeklemd, neemt de hepatische arteriële flow vrijwel onmiddellijk met ongeveer 30% toe (Jacab et al, 1995). Hoewel deze relatie vaak wordt beschreven als “semi-reciproque”, zoals in de meeste relaties één partner uiteindelijk al het werk doet; als de leverslagader wordt afgeklemd, doet de poortader niets om de doorstroming te verhogen.
Hoe gebeurt dit? De meest plausibele verklaring is de “adenosine washout hypothese”. Deze werd voorgesteld door Lautt e.a. (1985), en is in de literatuur blijven bestaan, ondanks het feit dat het bewijs ervoor nogal wankel is. Samengevat:
- Adenosine komt vrij in de ruimte van Mall, een periportale ruimte die wordt ingenomen door de poortader, de leverslagader en de galbuis.
- Het zit daar dan opgesloten, omdat de ruimte van Mall gescheiden is van andere vloeistofcompartimenten. Kortom, het kan nergens anders heen, dan diffunderen in de vaten om te worden weggespoeld.
- De poortader in deze ruimte heeft de hoogste stroomsnelheid, en daarom wordt, als de portale stroming snel is, veel van de adenosine uit de ruimte van Mall gespoeld.
- Aangezien adenosine een vasodilator is, leidt het verlies ervan tot vasoconstrictie.
- Aangezien de leverslagader het enige is in de ruimte van Mall met ook maar een greintje vasculaire gladde spier, wordt deze hierdoor het meest getroffen.
- Ergo, de portale doorstroming, door de hoeveelheid adenosine in de ruimte van Mall aan te passen, regelt de vasculaire weerstand van de leverslagader.
Dit idee schijnt het soort levensduur te hebben dat men zou verwachten van een theorie die in feite juist is, en de voornaamste uitdagers schijnen hoofdzakelijk de aard van de uitgewassen bemiddelaar te betwisten (d.w.z. sommigen beweren dat het stikstofmonoxide, ATP, koolmonoxide, enzovoort moet zijn). Omwille van het gezond verstand van de lezer zullen deze details worden overgelaten aan de kust waar ze werden gevonden.
Variabele zuurstofonttrekking door de lever
Uit alle voorgaande besprekingen zou men terecht kunnen concluderen dat, hoewel de bloedvoorziening van de lever duidelijk aan enige regulering onderhevig is, deze niet bijzonder nauw verbonden schijnt te zijn met de stofwisselingssnelheid ervan – zeker niet in dezelfde mate als bijvoorbeeld de cerebrale bloedsomloop verbonden is met het cerebrale metabolisme. Dit is redelijk juist. De belangrijkste reguleringsmechanismen, zoals de portale post-prandiale flowverhoging of de leverarteriële bufferrespons, zijn echt niet ontworpen om het aanbod op de vraag af te stemmen – zij lijken te zijn gericht op de
Dus moet de lever zich op andere manieren aanpassen aan de fluctuerende zuurstoftoevoer. Hij wijzigt namelijk zijn zuurstofonttrekkingsverhouding. Lutz e.a. (1975) ontdekten dat de relatie tussen zuurstofonttrekking en bloedstroom in wezen lineair was, d.w.z. dat naarmate de zuurstoftoevoer naar de lever afnam, de lever steeds meer zuurstof onttrok, totdat vrijwel alle zuurstof op was en het veneuze bloed van de lever zwart van de anoxie werd. Zoals duidelijk te zien is in dit diagram uit het oorspronkelijke artikel, neigt de extractieratio naar 100%.

Extrinsieke factoren die de perfusie van de lever beïnvloeden
De reden dat dit hier is opgenomen, is omdat de examinatoren in het collegecommentaar op vraag 13 uit het tweede paper van 2016 verwachtten dat een goed antwoord “draaide om de vraag hoe de bloedstroom in de lever wordt geregeld… …met betrekking tot intrinsieke en extrinsieke factoren”. Wat zijn die extrinsieke factoren? Als men nagaat hoe deze elders zijn weergegeven, komt men tot de conclusie dat de lijst onmogelijk breed kan zijn en factoren kan omvatten als “in de lever gestompt worden” en “dood van de bloedsomloop”. In plaats van deze te omschrijven als “controlemechanismen” of “regulerende factoren”, zou het eerlijker zijn ze te omschrijven als “invloeden van buitenaf die de leverdoorbloeding beïnvloeden, vaak op dramatische wijze, ondanks het feit dat de lever op de een of andere manier nog functioneert”. Om ze samen te vatten:
- Extrahepatische factoren die de hepatische perfusie verhogen:
- Verhoogde veneuze return
- Spontane ademhaling (inspiratie)
- Verhoogde arteriële bloedstroom
- Alles wat de cardiale output verhoogt
- Verhoogde portale bloedstroom
- Splanchnische vasodilatatie. b.v. na een maaltijd
- Verhoogde veneuze return
- Extrahepatische factoren die de leverperfusie verminderen
- Verlaagde veneuze return
- Positieve druk ventilatie
- Heartfalen, m.n. rechter hartfalen
- Fluid overload states, b.v. tussen regelmatige dialysesessies
- Verlaagde arteriële bloedstroom
- Alles wat de cardiale output vermindert, bv. hartfalen
- Alles wat de splanchnische bloedstroom herverdeelt, bv. lichaamsbeweging, vrijkomen van catecholamine, stress
- Verlaagde portale bloedstroom
- Splanchnische vasoconstrictie, bv. shocktoestanden
- Verlaagde veneuze return
Veranderingen in het medicijnmetabolisme als gevolg van veranderingen in de leverdoorbloeding
Vraag 13 uit het tweede paper van 2016 vroeg de cursisten ook om “de veranderingen in het medicijnmetabolisme te verklaren als de leverdoorbloeding afneemt”. Dit is eigenlijk een vraag over de leverklaring, die in detail wordt besproken in het onderdeel farmacokinetiek. Om het aantal kliks bij de examenrevisie te beperken, worden de belangrijke punten hier in de kortst mogelijke vorm weergegeven.
- De leverklaring is het product van de leverbloedstroom en de leverextractieratio:
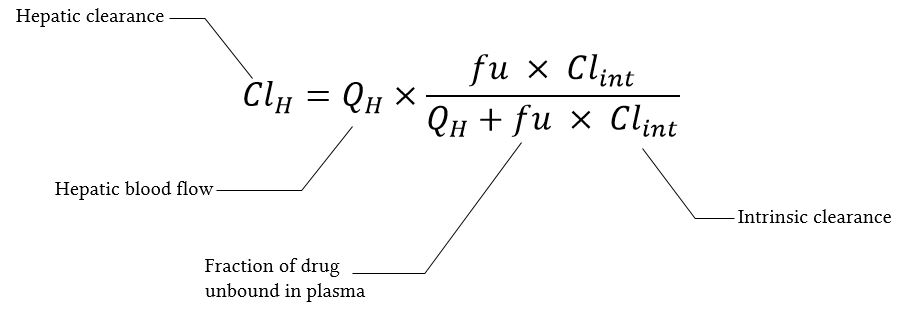
waarbij de leverextractieratio hier wordt weergegeven door alles buiten het “×”-symbool.
- De leverextractieratio is de fractie van het geneesmiddel dat in het bloed de lever binnenkomt en tijdens één passage van het bloed door de lever onomkeerbaar wordt verwijderd (geëxtraheerd).
-
Wanneer de leverbloedstroom afneemt, zal de leverextractieratio voor alle geneesmiddelen toenemen.
- Wat er met het metabolisme van een geneesmiddel gebeurt wanneer de leverbloedstroom afneemt, hangt af van de intrinsieke leverklaring van dat geneesmiddel.
- Hoe hoger de intrinsieke klaring, hoe meer de klaring van dat geneesmiddel afhankelijk is van de bloedstroom.
- Dus voor geneesmiddelen met een lage intrinsieke klaring zal de leverklaring niet significant toenemen bij toenemende bloedstroom.
- Voor geneesmiddelen met een hoge intrinsieke klaring zal de leverklaring tamelijk lineair afnemen, evenredig met de leverbloedstroom.
Geef een antwoord