Unterstamm Crustacea
Allgemeines über Krustentiere
Krustentiere unterscheiden sich von anderen Gliederfüßern in vielerlei Hinsicht, aber das einzige wirkliche Unterscheidungsmerkmal ist, dass Krustentiere die einzigen Gliederfüßer mit zwei Fühlerpaaren sind. Zusätzlich zu zwei Paar Fühlern und einem Paar Unterkiefer haben Krebstiere zwei Paar Oberkiefer am Kopf, gefolgt von einem Paar Anhängsel an jedem Körpersegment oder Somiten. Bei einigen Krebstieren tragen nicht alle Somiten Anhängsel. Alle Anhängsel, mit Ausnahme vielleicht der ersten Fühler, sind primitiv zweigliedrig (zwei Hauptäste), und zumindest einige Anhängsel heutiger erwachsener Tiere weisen diesen Zustand auf. Die auf die Atmung spezialisierten Organe fungieren, falls vorhanden, als Kiemen.
Die meisten Krebstiere haben zwischen 16 und 20 Somiten, aber einige Formen haben 60 Somiten oder mehr. Eine größere Anzahl von Somiten ist ein primitives Merkmal. Die abgeleitete Form hat weniger Segmente und eine stärkere Tagmatisierung. Die wichtigsten Tagmata sind Kopf, Thorax und Abdomen, aber diese sind nicht in der gesamten Klasse (oder sogar innerhalb einiger Unterklassen) homolog, da die Somiten unterschiedlich stark verschmolzen sind, wie z. B. beim Cephalothorax.
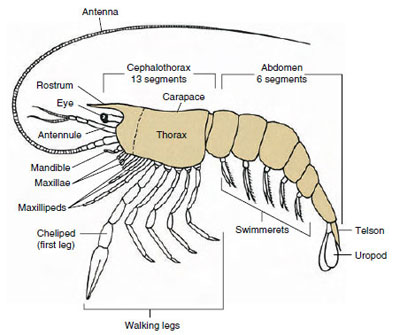
Abbildung 19-1
Archetypischer Grundriss der Malacostraca. Die beiden Maxillen und drei
Maxillipeden sind zur Veranschaulichung des
Grundrisses schematisch getrennt.
Die bei weitem größte Gruppe der Krebstiere ist die Klasse der Malacostraca, zu der Hummer, Krebse, Krabben, Strandläufer, Wanzen und viele andere gehören. Diese weisen eine erstaunlich konstante Anordnung von Körpersegmenten und Tagmata auf, die als Stammbaum der Klasse gilt (Abbildung 19-1). Dieser typische Körperbau besteht aus einem Kopf mit fünf (embryonal sechs) verschmolzenen Somiten, einem Thorax mit acht Somiten und einem Abdomen mit sechs Somiten (bei einigen wenigen Arten sieben). Am vorderen Ende befindet sich ein nicht segmentiertes Rostrum und am hinteren Ende ein nicht segmentiertes Telson, das zusammen mit dem letzten Abdomensomiten und seinen Uropoden bei vielen Formen einen Schwanzfächer bildet.
Bei vielen Krebstieren kann sich die dorsale Cuticula des Kopfes nach hinten und um die Seiten des Tieres herum erstrecken und einige oder alle Thorax- und Abdomensomiten bedecken oder mit ihnen verschmolzen sein. Diese Hülle wird als Panzer bezeichnet. Bei einigen Gruppen bildet der Panzer muschelartige Klappen, die den ganzen oder einen Großteil des Körpers bedecken. Bei den Zehnfußkrebsen (einschließlich Hummern, Garnelen, Krabben und anderen) bedeckt der Panzer den gesamten Cephalothorax, nicht aber den Hinterleib.
Form und Funktion
Aufgrund ihrer Größe und leichten Verfügbarkeit wurden große Krebstiere wie Krebse mehr als andere Gruppen untersucht. Sie werden auch häufig in einführenden Laborkursen untersucht. Daher gelten viele der folgenden Bemerkungen speziell für Krebse und ihre Verwandten.
Äußere Merkmale
Die Körper der Krebse sind mit einer sekretierten Kutikula bedeckt, die aus Chitin, Eiweiß und kalkhaltigem Material besteht. Die härteren, schweren Platten größerer Krebstiere sind besonders kalkhaltig. Die harte Schutzhülle ist an den Verbindungsstellen zwischen den Somiten weich und dünn, was eine flexible Bewegung ermöglicht. Der Panzer, sofern vorhanden, bedeckt einen Großteil oder den gesamten Cephalothorax; bei Zehnfußkrebsen wie den Krebsen sind alle Kopf- und Brustsegmente dorsal vom Panzer umgeben. Jeder Somit, der nicht vom Panzer umschlossen ist, wird von einer dorsalen Kutikularplatte, dem Tergum, bedeckt (Abbildung 19-2A), und ein ventraler Querbalken, das Sternum, liegt zwischen den Segmentanhängen (Abbildung 19-2B). Der Hinterleib endet in einem Telson, das nicht zu den Somiten gezählt wird und den Anus trägt. (Das Telson ist möglicherweise mit dem Pygidium der Ringelwürmer verwandt.)
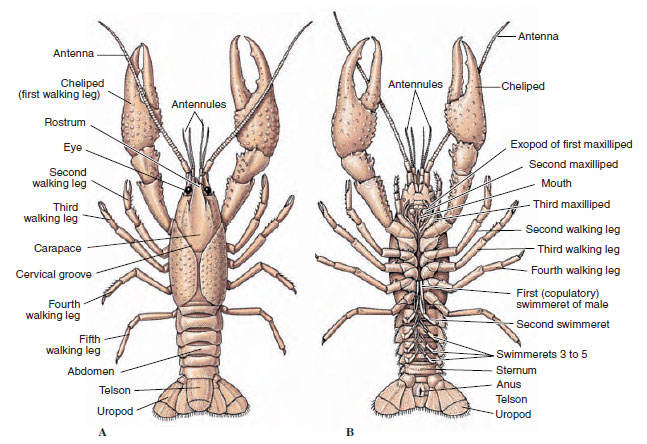
Abbildung 19-2
Außenbau der Krebse. A, Dorsalansicht. B, Ventralansicht.
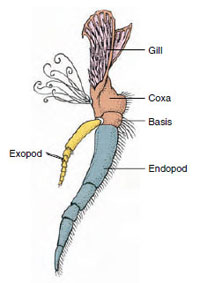
Abbildung 19-3
Teile eines zweigliedrigen Krebstieres
Anhangsgebilde (drittes Maxilliped eines
Krebses).
Die Position der Gonoporen variiert je nach Geschlecht und Gruppe der Krebse. Sie können sich auf oder an der Basis eines Anhängselpaares, am terminalen Ende des Körpers oder auf Somiten ohne Beine befinden. Bei den Krebsen befinden sich die Öffnungen der Samenleiter auf der medianen Seite an der Basis des fünften Schreitbeinpaars und die der Eileiter an der Basis des dritten Paars. Bei den Weibchen befindet sich die Öffnung des Samenbehälters gewöhnlich in der Mittellinie zwischen dem vierten und dem fünften Paar Schreitbeine.
Anhangsgebilde: Mitglieder der Klassen Malacostraca (einschließlich Krebse) und Remipedia haben typischerweise ein Paar gelenkige Anhänge an jedem Somiten (Abbildung 19-3), obwohl die Abdomensomiten in den anderen Klassen keine Anhänge tragen. Eine beträchtliche Spezialisierung ist bei den Anhängseln abgeleiteter Krebstiere wie den Flusskrebsen zu beobachten. Bei allen handelt es sich jedoch um Variationen des grundlegenden, zweigliedrigen Plans, der durch ein Krebsanhängsel wie ein Maxilliped (ein zu einem Kopfanhängsel umgebautes Thoraxglied) veranschaulicht wird (Abbildungen 19-3 und 19-4). Der basale Teil, die Protopode, trägt eine laterale Exopode und eine mediale Endopode. Die Protopode besteht aus einem oder zwei Gelenken (Basis und Coxa), während die Exopode und die Endopode jeweils ein bis mehrere Gelenke haben. Einige Anhängsel, wie z. B. die Laufbeine der Krebse, sind sekundär uniramidal geworden. An den Gliedmaßen von Krebstieren treten manchmal mediale oder laterale Fortsätze auf, die als Endit bzw. Exit bezeichnet werden, und ein Exit an der Protopode wird als Epipode bezeichnet. Epipoden sind oft als Kiemen modifiziert. Tabelle 19-1 zeigt, wie die verschiedenen Anhängsel vom biramischen Plan aus modifiziert wurden, um spezifische Funktionen zu erfüllen.
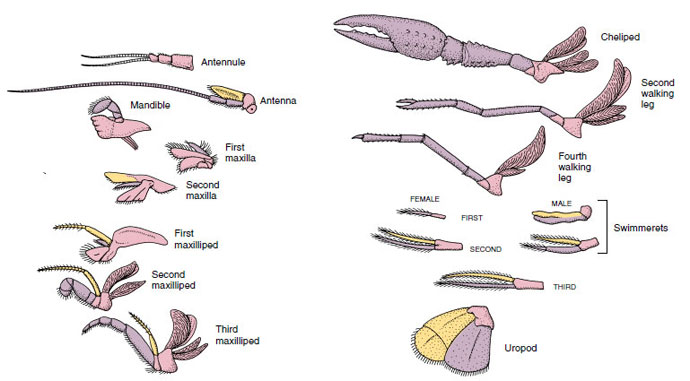
Abbildung 19-4
Anhangsgebilde eines Flusskrebses, die zeigen, wie sie vom biramösen Grundplan abgewandelt wurden, wie man sie bei einem
Schwimmkörper findet. Protopod, rosa; Endopod, violett; Exopod, gelb.
Strukturen, die einen ähnlichen Grundriss haben und von einer gemeinsamen Form abstammen, werden als homolog bezeichnet, unabhängig davon, ob sie die gleiche Funktion haben oder nicht. Da sich spezialisierte Laufbeine, Mundwerkzeuge, Fußsohlen und Schwimmfüße alle aus einem gemeinsamen biramischen Typ entwickelt haben, aber so modifiziert wurden, dass sie unterschiedliche Funktionen erfüllen, sind sie alle homolog zueinander, ein Zustand, der als serielle Homologie bekannt ist. Ursprünglich waren sich alle Gliedmaßen sehr ähnlich, aber im Laufe der Evolution wurden einige Zweige reduziert, einige gingen verloren, einige wurden stark verändert, und einige neue Teile kamen hinzu. Krebse und ihre Verbündeten besitzen die ausgefeilteste serielle Homologie im Tierreich, sie haben 17 verschiedene, aber seriell homologe Arten von Anhängseln (Tabelle 19-1).
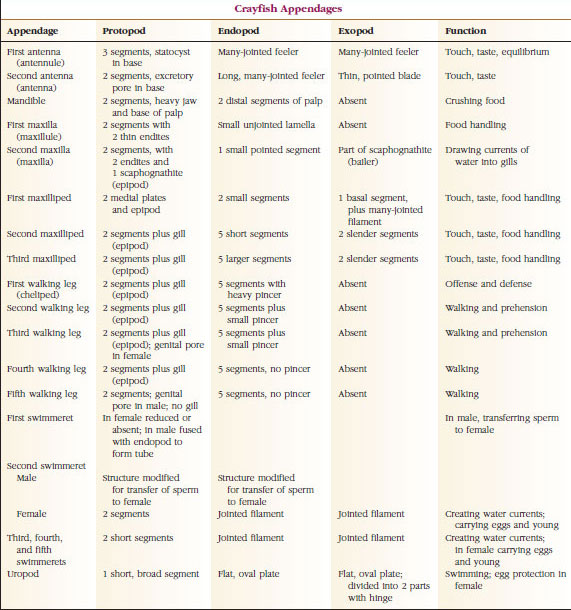
TABELLE 19.1
Interne Merkmale
Das Muskel- und Nervensystem sowie die Segmentierung im Thorax und im Abdomen zeigen deutlich den Metamerismus, der von annelidenähnlichen Vorfahren geerbt wurde, aber es gibt deutliche Modifikationen in anderen Systemen. Bei den meisten Veränderungen handelt es sich um eine Konzentration von Teilen in einer bestimmten Region oder um die Verringerung oder den vollständigen Verlust von Teilen, wie z. B. der Intersepta.
Hämocoel: Der wichtigste Körperraum der Arthropoden ist nicht das Coelom, sondern ein blutgefülltes Hemocoel. Während der Embryonalentwicklung der meisten Gliederfüßer öffnen sich rudimentäre Coelomhöhlen innerhalb des Mesoderms zumindest einiger Somiten. Diese werden bald verödet oder gehen in den Raum zwischen den sich entwickelnden mesodermalen und ektodermalen Strukturen und dem Dotter über. Dieser Raum wird zum Hemocoel und ist somit nicht von einem mesodermalen Peritoneum ausgekleidet. Bei Krebstieren sind die einzigen verbliebenen Coelomkompartimente die Endsäcke der Ausscheidungsorgane und der Raum um die Gonaden.
Muskulatur: Gestreifte Muskeln machen einen beträchtlichen Teil des Körpers der meisten Crustacea aus. Die Muskeln sind in der Regel in antagonistischen Gruppen angeordnet: Beuger, die einen Teil zum Körper ziehen, und Strecker, die ihn nach außen ziehen. Der Bauch eines Flusskrebses hat kräftige Beugemuskeln (Abbildung 19-5), die benutzt werden, wenn das Tier rückwärts schwimmt – sein bestes Mittel zur Flucht. Kräftige Muskeln auf beiden Seiten des Magens steuern die Unterkiefer.
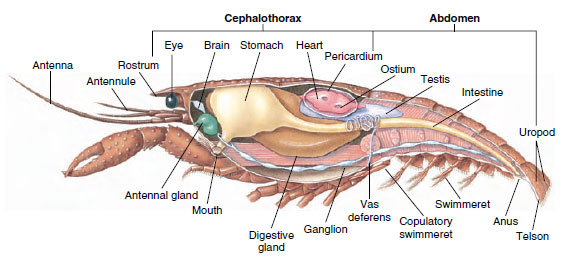
Abbildung 19-5
Interner Aufbau eines männlichen Flusskrebses.
Atmungssystem: Der Atemgasaustausch bei kleineren Krebstieren erfolgt über dünnere Bereiche der Cuticula (z. B. in den Beinen) oder den gesamten Körper, und spezialisierte Strukturen können fehlen. Größere Krebstiere haben Kiemen, bei denen es sich um zarte, federartige Fortsätze mit sehr dünner Cuticula handelt. Bei Zehnfußkrebsen umschließen die Seiten des Panzers die Kiemenhöhle, die nach vorne und nach ventral offen ist (Abbildung 19-6). Kiemen können von der Pleurawand in die Kiemenhöhle hineinragen, von der Verbindung der Thoraxbeine mit dem Körper oder von den Thoraxcoxen ausgehen. Die beiden letztgenannten Arten sind typisch für Krebse. Der „Schöpfer“, ein Teil des zweiten Oberkiefers, zieht das Wasser über die Kiemenfäden in die Kiemenhöhle an den Basen der Beine und aus der Kiemenhöhle an der Vorderseite heraus.
Kreislaufsystem: Krustentiere und andere Gliederfüßer haben ein „offenes“ oder lakunares Kreislaufsystem. Das bedeutet, dass es keine Venen gibt und keine Trennung von Blut und Zwischenzellflüssigkeit wie bei Tieren mit geschlossenem System. Die Hämolymphe (das Blut) verlässt das Herz über die Arterien, zirkuliert durch das Hämocoel und kehrt in die venösen Sinus oder Räume anstelle der Venen zurück, bevor sie wieder in das Herz eintritt. Anneliden haben ein geschlossenes System, ebenso wie die Wirbeltiere.
Das Herz ist das Hauptantriebsorgan. Es ist ein einkammeriger Sack aus quergestreiften Muskeln. Die Hämolymphe tritt aus dem umgebenden Sinus pericardialis durch paarige Ostien in das Herz ein, wobei Klappen den Rückfluss in den Sinus verhindern (Abbildung 19-6). Vom Herzen aus gelangt die Hämolymphe in eine oder mehrere Arterien. Ventile in den Arterien verhindern einen Rückfluss der Hämolymphe. Kleine Arterien entleeren sich in Gewebesinus, die wiederum oft in einen großen Sinus sternalis münden (Abbildung 19-6).
Von dort aus leiten afferente Sinuskanäle die Hämolymphe zu den Kiemen, falls vorhanden, um Sauerstoff und Kohlendioxid auszutauschen. Die Hämolymphe kehrt dann durch efferente Kanäle zum Sinus pericardialis zurück (Abbildung 19-6).
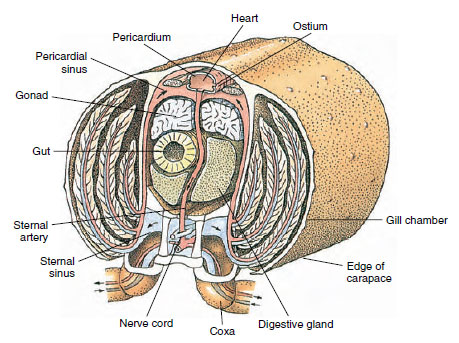
Abbildung 19-6
Diagrammatischer Querschnitt durch die Herzregion eines Flusskrebses, der die Richtung des Blutflusses in diesem „offenen“
Blutsystem zeigt. Das Herz pumpt das Blut durch die Arterien zu den Körpergeweben, die in die Gewebesäulen münden. Das zurückfließende
Blut gelangt in den Sinus sternalis, dann durch die Kiemen zum Gasaustausch und schließlich über ableitende
Kanäle zurück in den Sinus pericardialis. Man beachte das Fehlen von Venen.
Die Hämolymphe der Gliederfüßer ist weitgehend farblos. Sie enthält amöboide Zellen von mindestens zwei Typen. Hämocyanin, ein kupferhaltiges Atmungspigment, oder Hämoglobin, ein eisenhaltiges Pigment, können in Lösung getragen werden. Hämolymphe hat die Eigenschaft, zu gerinnen, was ihren Verlust bei kleineren Verletzungen verhindert. Einige ameboide Zellen setzen einen thrombinähnlichen Gerinnungsstoff frei, der die Gerinnung verhindert.
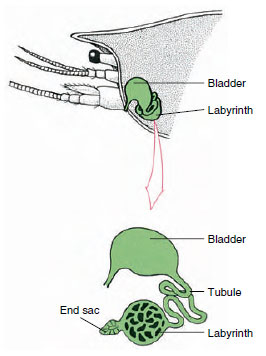
Abbildung 19-7
Schema der Antennendrüse (grüne Drüse) der
Krebse. (In natürlicher Lage ist das Organ stark gefaltet.) Einigen Krebstieren fehlt ein Labyrinth,
und der Ausscheidungstubus (Nephridialkanal) ist
ein stark gewundenes Rohr.
Ausscheidungssystem: Die Ausscheidungsorgane erwachsener Krebstiere bestehen aus einem Paar röhrenförmiger Strukturen, die sich im ventralen Teil des Kopfes vor der Speiseröhre befinden (Abbildung 19-5). Sie werden als Antennendrüsen oder Maxillardrüsen bezeichnet, je nachdem, ob sie an der Basis der Fühler oder der zweiten Maxillae münden. Einige wenige erwachsene Krebstiere haben beides. Die Ausscheidungsorgane der Zehnfußkrebse sind die Antennendrüsen, die in dieser Gruppe auch grüne Drüsen genannt werden. Krebstiere haben keine malpighischen Tubuli, die Ausscheidungsorgane von Spinnen und Insekten.
Der Endsack der Antennendrüse, der aus einem embryonalen Coelomkompartiment hervorgeht, besteht aus einem kleinen Bläschen (Sacculus) und einer schwammartigen Masse, die Labyrinth genannt wird. Das Labyrinth ist über einen Ausscheidungstubus mit einer dorsalen Blase verbunden, die sich durch eine Pore an der ventralen Oberfläche des basalen Antennensegments nach außen hin öffnet (Abbildung 19-7). Der hydrostatische Druck innerhalb des Hämocoels sorgt für die Filtration der Flüssigkeit in den Endsack. Während das Filtrat den Ausscheidungstubus und die Blase passiert, wird es durch die Resorption von Salzen, Aminosäuren, Glukose und etwas Wasser modifiziert und schließlich als Urin ausgeschieden.
Die Ausscheidung von stickstoffhaltigen Abfällen (hauptsächlich Ammoniak) erfolgt durch Diffusion durch dünne Bereiche der Kutikula, insbesondere durch die Kiemen, und die so genannten Ausscheidungsorgane haben hauptsächlich die Aufgabe, die ionische und osmotische Zusammensetzung der Körperflüssigkeiten zu regulieren. Süßwasserkrebse, wie z. B. Krebse, sind ständig von einer Überverdünnung ihres Blutes durch Wasser bedroht, das über die Kiemen und andere wasserdurchlässige Oberflächen diffundiert. Die grünen Drüsen, die einen verdünnten, salzarmen Urin bilden, wirken als wirksame „Flutkontrolle“. Ein Teil von Na+ und Cl- geht mit dem Urin verloren, aber dieser Verlust wird durch die aktive Absorption von gelöstem Salz durch die Kiemen kompensiert. Bei marinen Krustentieren wie Hummern und Krabben dient die Niere dazu, die Salzzusammensetzung der Hämolymphe durch selektive Veränderung des Salzgehalts des Röhrenurins anzupassen. Bei diesen Formen bleibt der Urin gegenüber dem Blut isosmotisch.
Nervensystem und sensorische Systeme: Das Nervensystem der Krebstiere und der Ringelwürmer hat viele Gemeinsamkeiten, obwohl bei den Krebstieren die Ganglien stärker verschmolzen sind (Abbildung 19-5). Das Gehirn besteht aus einem Paar supraösophagealer Ganglien, die Nerven zu den Augen und zwei Fühlerpaaren führen. Es ist durch Bindeglieder mit dem subösophagealen Ganglion verbunden, einem Zusammenschluss von mindestens fünf Ganglienpaaren, die den Mund, die Fortsätze, die Speiseröhre und die Antennendrüsen mit Nerven versorgen. Der doppelte ventrale Nervenstrang hat ein Ganglienpaar für jeden Somiten und Nerven, die die Anhänge, Muskeln und andere Teile versorgen.
Zusätzlich zu diesem zentralen System kann es ein sympathisches Nervensystem geben, das mit dem Verdauungstrakt verbunden ist.
Krebse haben besser entwickelte Sinnesorgane als Anneliden. Die größten Sinnesorgane der Krebse sind die Augen und Statocysten. Über den ganzen Körper verteilt befinden sich Tasthaare, feine Ausstülpungen der Cuticula, die besonders häufig an den Chelizeren, den Mundwerkzeugen und dem Telson zu finden sind. Die chemischen Geschmacks- und Geruchssinne befinden sich in Haaren auf den Antennen, den Mundwerkzeugen und an anderen Stellen.
Eine sackartige Statozyste, die sich durch eine dorsale Pore zur Oberfläche hin öffnet, befindet sich auf dem Basalsegment jeder ersten Antenne der Krebse. Die Statozyste enthält einen Grat, der Sinneshaare aus der Chitinauskleidung und Sandkörner trägt, die als Statolithen dienen. Wann immer das Tier seine Position verändert, werden entsprechende Positionsänderungen der Körner auf den Sinneshaaren als Reize an das Gehirn weitergeleitet, und das Tier kann sich entsprechend anpassen. Jede Häutung (Ekdysis) führt zum Verlust der Kutikularauskleidung der Statozyste und damit der Sandkörner. Nach der Ekdysis werden neue Körner durch die Dorsalpore aufgenommen.
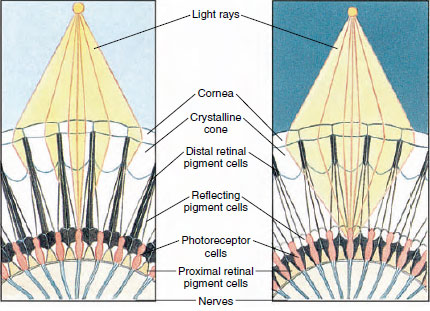
Abbildung 19-8
Ausschnitt aus dem Facettenauge eines Arthropoden, der die Pigmentwanderung
in den Ommatidien für das Tag- und Nachtsehen zeigt. Fünf Ommatidien sind in
jeder Abbildung dargestellt. Am Tag ist jedes Ommatidium von einem dunklen
Pigmentkragen umgeben, so dass jedes Ommatidium nur durch Lichtstrahlen
gereizt wird, die in seine eigene Hornhaut eindringen (Mosaiksehen); in der Nacht bildet das Pigment
unvollständige Kragen und die Lichtstrahlen können sich auf benachbarte Ommatidien
ausbreiten (kontinuierliches oder überlagerndes Bild).
Die Augen vieler Krebstiere sind zusammengesetzt und bestehen aus vielen Photorezeptoreinheiten, die Ommatidien genannt werden (Abbildung 19-8). Die abgerundete Oberfläche jedes Auges ist von einer durchsichtigen Kutikula, der Hornhaut, bedeckt, die in viele kleine Quadrate oder Sechsecke unterteilt ist, die als Facetten bezeichnet werden. Diese Facetten sind die äußeren Enden der Ommatidien. Jedes Ommatidium verhält sich wie ein winziges Auge und enthält mehrere Arten von Zellen, die säulenförmig angeordnet sind (Abbildung 19-8). Schwarze Pigmentzellen befinden sich zwischen benachbarten Ommatidien.
Die Bewegung der Pigmente in einem Gliederfüßer-Auge ermöglicht es ihm, sich an unterschiedliche Lichtmengen anzupassen. In jedem Ommatidium gibt es drei Gruppen von Pigmentzellen: distale retinale, proximale retinale und reflektierende; diese sind so angeordnet, dass sie einen mehr oder weniger vollständigen Kragen oder eine Hülle um jedes Ommatidium bilden können. Bei starkem Licht oder Tagesanpassung bewegt sich das distale Netzhautpigment nach innen und trifft auf das sich nach außen bewegende proximale Netzhautpigment, so dass sich eine vollständige Pigmenthülle um das Ommatidium bildet (Abbildung 19-8). In diesem Zustand erreichen nur Strahlen, die direkt auf die Hornhaut treffen, die Photorezeptorzellen (Netzhautzellen), da jedes Ommatidium von den anderen abgeschirmt ist. Somit sieht jedes Ommatidium nur einen begrenzten Bereich des Gesichtsfeldes (ein Mosaik- oder Appositionsbild). Bei schwachem Licht trennen sich distale und proximale Pigmente, so dass die Lichtstrahlen mit Hilfe der reflektierenden Pigmentzellen die Möglichkeit haben, sich auf benachbarte Ommatidien auszubreiten und ein kontinuierliches oder überlagerndes Bild zu bilden. Diese zweite Art des Sehens ist weniger präzise, nutzt aber die begrenzte Menge des empfangenen Lichts optimal aus.
Reproduktion, Lebenszyklen und endokrine Funktion
Die meisten Krebstiere haben getrennte Geschlechter, und es gibt verschiedene Spezialisierungen für die Kopulation bei verschiedenen Gruppen. Seepocken sind einhäusig, praktizieren aber im Allgemeinen Fremdbefruchtung. Bei einigen Ostracoden gibt es nur wenige Männchen, und die Fortpflanzung erfolgt meist parthenogenetisch. Die meisten Krebstiere brüten ihre Eier auf irgendeine Weise aus: Branchiopoden und Seepocken haben spezielle Brutkammern, Copepoden haben Brutsäcke, die an den Seiten des Abdomens befestigt sind (siehe Abbildung 19-19), und viele Malakostraken tragen Eier und Jungtiere an ihren Hinterleibsanhängseln.
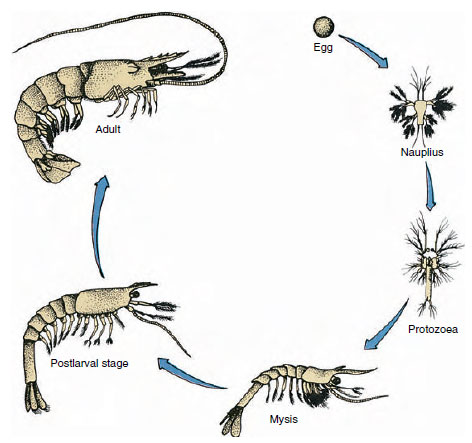
Abbildung 19-9
Lebenszyklus der Golfgarnele Penaeus. Penaeiden laichen in Tiefen von 40 bis 90 m. Die jungen Larvenformen sind
planktonisch und wandern küstennah in Gewässer mit geringerem Salzgehalt, um sich als Jungtiere zu entwickeln. Ältere Garnelen kehren in tieferes Wasser
offshore zurück.
Krebse haben eine direkte Entwicklung: es gibt keine Larvenform. Aus dem Ei schlüpft ein winziges Jungtier, das die gleiche Form wie das erwachsene Tier hat und über einen vollständigen Satz von Anhängseln und Somiten verfügt. Bei den meisten Krebstieren verläuft die Entwicklung jedoch indirekt, und aus dem Ei schlüpft eine Larve, die dem Erwachsenen in Struktur und Aussehen sehr ähnlich ist. Der Übergang von der Larve zum erwachsenen Tier ist die Metamorphose. Die primitivste und am häufigsten vorkommende Larve bei den Krebstieren ist die Nauplie (Abbildung 19-9 und 19-23). Nauplien tragen nur drei Anhängselpaare: uniramide erste Antennen, biramide Fühler und biramide Mandibeln. Alle fungieren in diesem Stadium als schwimmende Anhängsel.
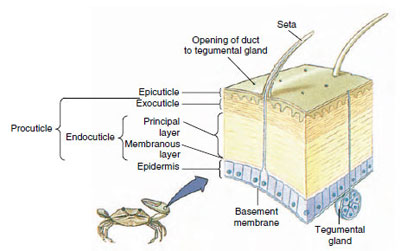
Abbildung 19-10
Struktur der Kutikula von Krebstieren.
Die nachfolgende Entwicklung kann eine allmähliche Veränderung zur erwachsenen Körperform beinhalten, und Anhänge und Somiten werden durch eine Reihe von Häutungen hinzugefügt, oder die Annahme der erwachsenen Form kann mit abrupteren Veränderungen einhergehen. Zum Beispiel verläuft die Metamorphose einer Seepocke von einer freischwimmenden Nauplie zu einer Larve mit einem muschelartigen Panzer, genannt Cypride, und schließlich zu einem sessilen Erwachsenen mit Kalkplatten.
Ecdysis: Die Ekdysis (ek’duh-sis) (gr. ekdyein, abstreifen) oder Häutung ist notwendig, damit der Körper an Größe zunimmt, da das Exoskelett nicht lebendig ist und nicht mitwächst, wenn das Tier wächst. Ein Großteil der Funktionsweise eines Krustentiers, einschließlich seiner Fortpflanzung, seines Verhaltens und vieler Stoffwechselvorgänge, wird direkt von der Physiologie des Häutungszyklus beeinflusst.
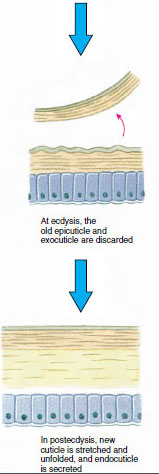
Das Häutchen, das von der darunter liegenden Epidermis abgesondert wird, hat mehrere Schichten (Abbildung 19-10). Die äußerste ist die Epikutikula, eine sehr dünne Schicht aus mit Lipiden imprägniertem Protein. Der Hauptteil der Cuticula besteht aus mehreren Schichten der Procuticula: (1) die Exokutikula, die direkt unter der Epikutikula liegt und Eiweiß, Kalziumsalze und Chitin enthält; (2) die Endokutikula, die ihrerseits aus (3) einer Hauptschicht, die mehr Chitin und weniger Eiweiß enthält und stark verkalkt ist, und (4) einer nicht verkalkten Membranschicht, einer relativ dünnen Schicht aus Chitin und Eiweiß, besteht.
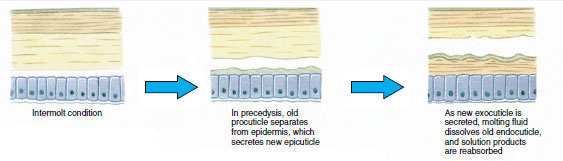
Einige Zeit vor der eigentlichen Ekdysis vergrößern sich die Epidermiszellen erheblich. Sie lösen sich von der Membranschicht, scheiden eine neue Epikutikula aus und beginnen mit der Absonderung einer neuen Exokutikula (Abbildung 19-11). In den Bereich oberhalb der neuen Epikutikula werden Enzyme freigesetzt. Diese Enzyme beginnen, die alte Endocuticula aufzulösen, und die löslichen Produkte werden resorbiert und im Körper des Krustentiers gespeichert. Einige Kalziumsalze werden als Gastrolithen (mineralische Ablagerungen) in den Magenwänden gespeichert. Schließlich bleiben von der alten Cuticula nur noch Exocuticula und Epicuticula übrig, die von neuer Epicuticula und neuer Exocuticula überlagert werden. Das Tier schluckt Wasser, das es über den Darm aufnimmt, und sein Blutvolumen nimmt stark zu. Durch den inneren Druck wird die Cuticula gespalten, und das Tier zieht sich aus seinem alten Exoskelett heraus (Abbildung 19-12). Es folgen eine Dehnung der noch weichen neuen Cuticula, die Ablagerung der neuen Endocuticula, die erneute Ablagerung der geretteten anorganischen Salze und anderer Bestandteile sowie die Aushärtung der neuen Cuticula. Während der Häutung ist das Tier wehrlos und bleibt im Verborgenen.
 |
|
 |
 |
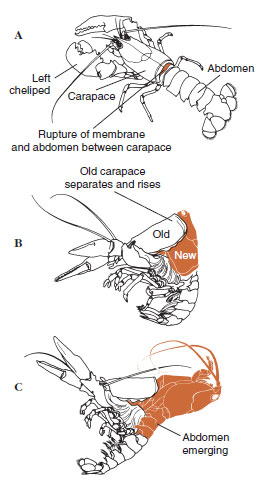
| Abbildung 19-12 Häutungssequenz bei einem Hummer, Homarus americanus. A: Die Membran zwischen Panzer und Hinterleib reißt, und der Panzer hebt sich langsam an. Dieser Schritt kann bis zu 2 Stunden dauern. B und C: Kopf, Thorax und schließlich Abdomen ziehen sich zurück. Dieser Vorgang dauert normalerweise nicht länger als 15 Minuten. Unmittelbar nach der Ekdysis sind die Pfoten ausgetrocknet und der Körper ist sehr weich. Der Hummer nimmt weiterhin schnell Wasser auf, so dass der Körper innerhalb von 12 Stunden um etwa 20 % in der Länge und 50 % im Gewicht zunimmt. Das Gewebewasser wird in den folgenden Wochen durch Proteine ersetzt. |
Abbildung 19-11 Sekretion und Resorption der Häutchen bei der Ekdysis. |
Wenn ein Krustentier jung ist, muss die Ekdysis häufig stattfinden, um das Wachstum zu ermöglichen, und der Häutungszyklus ist relativ kurz. Wenn sich das Tier der Geschlechtsreife nähert, werden die Häutungsintervalle immer länger, und bei einigen Arten hört die Häutung ganz auf. Während der Häutungsperioden kommt es zu einer Zunahme der Gewebemasse, da lebendes Gewebe Wasser ersetzt.
Hormonelle Steuerung des Ekdysis-Zyklus
Obwohl die Ekdysis hormonell gesteuert wird, wird der Zyklus häufig durch einen Umweltreiz ausgelöst, der vom zentralen Nervensystem wahrgenommen wird. Solche Reize können die Temperatur, die Tageslänge und die Luftfeuchtigkeit (bei Landkrabben) sein. Das Signal des Zentralnervensystems verringert die Produktion eines die Häutung hemmenden Hormons durch das X-Organ. Das X-Organ ist eine Gruppe von neurosekretorischen Zellen in der Medulla terminalis des Gehirns. Bei Flusskrebsen und anderen Zehnfußkrebsen befindet sich die Medulla terminalis im Augenstiel. Das Hormon wird in den Axonen des X-Organs zur Sinusdrüse (die selbst wahrscheinlich keine Drüsenfunktion hat), ebenfalls im Augenstiel, transportiert, wo es in die Hämolymphe freigesetzt wird.
Ein Absinken des Spiegels des Häutungshormons fördert die Ausschüttung eines Häutungshormons aus den Y-Organen. Die Y-Organe befinden sich unter der Epidermis in der Nähe der Adduktorenmuskeln der Mandibeln und sind mit den Prothoraxdrüsen der Insekten verwandt, die das Hormon Ecdyson produzieren. Die Wirkung des Häutungshormons besteht darin, die Prozesse einzuleiten, die zur Ekdysis (Proökdysis) führen. Sobald der Zyklus eingeleitet ist, läuft er automatisch ab, ohne dass weitere Hormone aus den X- oder Y-Organen einwirken.
Weitere endokrine Funktionen: Die Entfernung der Augenstiele beschleunigt nicht nur die Häutung, sondern es wurde auch vor über 100 Jahren festgestellt, dass Krebse, denen die Augenstiele entfernt wurden, ihre Körperfärbung nicht mehr an die Hintergrundbedingungen anpassen können. Vor über 50 Jahren entdeckte man, dass dieser Fehler nicht durch den Verlust des Sehvermögens, sondern durch den Verlust von Hormonen in den Augenstielen verursacht wurde. Die Körperfarbe der Krebstiere wird größtenteils von Pigmenten in speziellen verzweigten Zellen (Chromatophoren) in der Epidermis bestimmt.
Die Konzentration der Pigmentkörnchen im Zentrum der Zellen bewirkt eine Aufhellung, die Verteilung des Pigments in den Zellen eine Abdunkelung. Das Pigmentverhalten wird durch Hormone aus neurosekretorischen Zellen im Augenstiel gesteuert, ebenso wie die Migration von Netzhautpigmenten für die Hell-Dunkel-Anpassung in den Augen (Abbildung 19-8).
Die Freisetzung von neurosekretorischem Material aus den Perikardorganen in der Wand des Herzbeutels bewirkt eine Erhöhung der Herzschlagrate und -amplitude.
Androgene Drüsen, die erstmals bei einem Amphipoden (Orchestia, ein gewöhnlicher Strandhüpfer) gefunden wurden, kommen bei männlichen Malakostracern vor. Im Gegensatz zu den meisten anderen endokrinen Organen der Krebstiere handelt es sich nicht um neurosekretorische Organe. Ihre Sekretion stimuliert die Ausprägung der männlichen Geschlechtsmerkmale. Junge Malakostraken haben rudimentäre androgene Drüsen, aber bei den Weibchen entwickeln sich diese Drüsen nicht. Wenn sie einem Weibchen künstlich eingepflanzt werden, verwandeln sich ihre Eierstöcke in Hoden und beginnen, Spermien zu produzieren, und ihre Anhängsel nehmen bei der nächsten Häutung männliche Merkmale an. Bei den Asseln befinden sich die androgenen Drüsen in den Hoden; bei allen anderen Malakostracern liegen sie zwischen den Muskeln der Coxopoden der letzten Thoraxbeine und sind teilweise in der Nähe der Enden der Vas deferentia angebracht. Obwohl die Weibchen keine androgenähnlichen Organe besitzen, produzieren ihre Eierstöcke ein oder zwei Hormone, die die sekundären Geschlechtsmerkmale beeinflussen.
Hormone, die andere Körperprozesse bei Crustacea beeinflussen, können vorhanden sein, und es gibt Hinweise darauf, dass eine neurosekretorische Substanz, die im Augenstiel produziert wird, den Blutzuckerspiegel reguliert.
Futtergewohnheiten
Futtergewohnheiten und Anpassungen bei der Nahrungsaufnahme variieren stark zwischen den Krebstieren. Viele Formen können je nach Umgebung und Nahrungsangebot von einer Art der Nahrungsaufnahme zur anderen wechseln, aber alle verwenden die gleichen grundlegenden Mundwerkzeuge. Unterkiefer und Oberkiefer dienen der Nahrungsaufnahme; die Oberkiefer halten und zerkleinern die Nahrung. Bei Raubtieren dienen die Laufbeine, insbesondere die Chelicipeden, dem Nahrungserwerb.
Viele Krebstiere, sowohl große als auch kleine, sind räuberisch, und einige haben interessante Anpassungen zum Töten der Beute. Eine garnelenähnliche Form, Lygiosquilla, hat an einem ihrer Laufbeine einen spezialisierten Finger, der in eine Rille gezogen und plötzlich losgelassen werden kann, um eine vorbeigehende Beute zu durchbohren. Pistolenkrebse (Alpheus spp.) haben eine enorm vergrößerte Chela, die wie der Hammer eines Gewehrs gespannt werden kann und mit einer Kraft zuschlägt, die ihre Beute betäubt.
Die Nahrung von Schwebfressern reicht von Plankton und Detritus bis hin zu Bakterien. Räuber verzehren Larven, Würmer, Krebstiere, Schnecken und Fische. Aasfresser ernähren sich von toten tierischen und pflanzlichen Stoffen. Suspensionsfresser wie Flohkrebse, Wasserflöhe und Seepocken benutzen ihre Beine, die einen dicken Saum aus Seten tragen, um Wasserströmungen zu erzeugen, die Nahrungspartikel durch die Seten hindurchfegen. Schlammgarnelen (Upogebia spp.) verwenden lange Seten an den ersten beiden Paaren ihrer Brustanhänge, um durch die Bewegungen ihrer Schwimmfüße Nahrungsmaterial aus dem Wasser zu filtern, das durch ihre Höhle zirkuliert.
Krebse haben einen zweiteiligen Magen (Abbildung 19-13). Der erste Teil enthält eine Magenmühle, in der die bereits von den Mandibeln zerkleinerte Nahrung durch drei Kalkzähne weiter in Partikel zermahlen werden kann, die fein genug sind, um durch einen Setosefilter im zweiten Teil zu gelangen; die Nahrungspartikel gelangen dann zur chemischen Verdauung in den Darm.
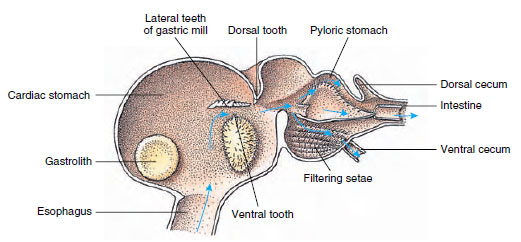
Abbildung 19-13
Malacostracan-Magen mit Magen-„Mühle“ und Richtung der Nahrungsbewegungen. Die Mühle hat Chitinkämme oder
Zähne zum Kauen und Seten, um die Nahrung zu zerdrücken, bevor sie in den Pylorusmagen gelangt.
Schreibe einen Kommentar