Neurotransmitterfreisetzung
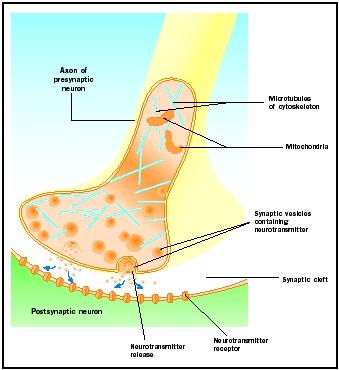
Aktionspotentiale, die an synaptischen Knoten ankommen, lösen die Freisetzung von Neurotransmitter in den synaptischen Spalt aus. Der molekulare Mechanismus ist noch nicht vollständig geklärt. Zwischen dem Eintreffen des Aktionspotentials und der Freisetzung von Neurotransmittern tritt eine „synaptische Verzögerung“ von ein bis zwei Millisekunden auf. Aktionspotenziale öffnen Kalziumkanäle in der Membran des synaptischen Knorpels, was eine Einwärtsbewegung von Kalziumionen bewirkt. Die Kalziumionen lösen die Freisetzung von Neurotransmitter aus den synaptischen Vesikeln in den synaptischen Spalt aus. Bei diesem Prozess der Exozytose verschmelzen die synaptischen Vesikel mit der präsynaptischen Membran. Die Membranen der alten Vesikel werden Teil der präsynaptischen Membran, und neue Vesikel werden von einem benachbarten Membranbereich abgezwickt. Diese neuen Vesikel werden anschließend mit neu synthetisierten oder „recycelten“ Neurotransmittern wieder aufgefüllt.
Die freigesetzten Neurotransmitter diffundieren durch den engen synaptischen Spalt. An der postsynaptischen Membran binden Neurotransmittermoleküle an membrangebundene Rezeptormoleküle mit für den jeweiligen Neurotransmitter spezifischen Erkennungsstellen. Die Bindung des Neurotransmitters an den Rezeptor löst eine für diesen Rezeptor spezifische postsynaptische Reaktion aus. Diese Reaktionen können je nach den Eigenschaften des Rezeptors entweder erregend oder hemmend sein. Wenn die Rezeptorstimulation dazu führt, dass die postsynaptische Membran elektrisch positiver wird (depolarisiert), handelt es sich um ein exzitatorisches postsynaptisches Potenzial (EPSP). Wird es negativer (hyperpolarisiert), handelt es sich um ein inhibitorisches postsynaptisches Potenzial (IPSP). Erregung und Hemmung hängen von den Eigenschaften des Rezeptors und nicht des Neurotransmitters ab. An Natrium- oder Kalziumkanäle gekoppelte Rezeptoren sind erregend und bewirken eine Depolarisation der postsynaptischen Membran, während an Chlorid- oder Kaliumkanäle gekoppelte Rezeptoren hemmend wirken und eine Hyperpolarisation der postsynaptischen Membran bewirken. Solche an Ionenkanäle gekoppelten Rezeptoren werden als ionotrope Rezeptoren bezeichnet.
Andere Rezeptoren sind an „Second-Messenger“-Systeme gekoppelt, die eine Reihe von biochemischen Reaktionen in der postsynaptischen Zelle auslösen. Dies sind metabotrope Rezeptoren. Metabotrope Rezeptoren können viele verschiedene
Postsynaptische Ereignisse. Diese reichen von der direkten Aktivierung benachbarter Ionenkanäle über die Veränderung der Rezeptorempfindlichkeit bis hin zur Transkription spezifischer Boten-Ribonukleinsäuren (RNAs) oder sogar der Aktivierung spezifischer Gene . Chemische Synapsen sind Teil eines sehr anpassungsfähigen und flexiblen Kommunikationssystems. Es handelt sich nicht um statische anatomische Strukturen mit festen Eigenschaften, sondern um dynamische Strukturen, die ihre molekularen Eigenschaften je nach den sich ändernden Umständen ändern können.
Es gibt buchstäblich Hunderte von Neurotransmittern. Einige sind recht einfache Verbindungen wie Acetylcholin, Serotonin, die Katecholamine (Dopamin, Noradrenalin und Epinephrin) und eine Reihe von Aminosäuren. Viele von ihnen sind komplexer und gehören zur großen Gruppe der Neuropeptid-Transmitter. Einmal in den synaptischen Spalt freigesetzt, bleiben Neurotransmitter so lange aktiv, bis sie entweder chemisch verändert oder durch spezielle Trägersysteme in den synaptischen Knoten zurückgebracht und recycelt werden. An cholinergen Synapsen befindet sich im synaptischen Spalt Acetylcholinesterase. Dieses Enzym spaltet den Neurotransmitter in Acetat und Cholin, von denen keines aktiv ist. Serotonin und Epinephrin hingegen werden in das präsynaptische Terminal aufgenommen und recycelt.
Schreibe einen Kommentar