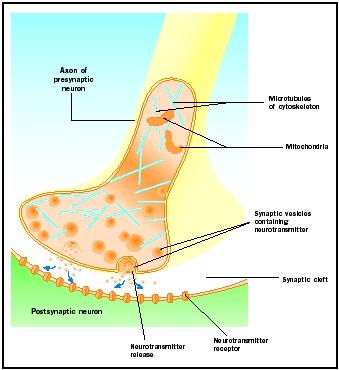
Eliberarea neurotransmițătorilor
Potențialele de acțiune care ajung la butoanele sinaptice declanșează eliberarea de neurotransmițător în fanta sinaptică. Mecanismul molecular nu este complet înțeles. Între sosirea potențialului de acțiune și eliberarea neurotransmițătorului are loc o „întârziere sinaptică” de una până la două milisecunde. Potențialele de acțiune deschid canalele de calciu în membrana butonului sinaptic, ceea ce determină o mișcare interioară a ionilor de calciu . Ionii de calciu declanșează eliberarea de neurotransmițător din veziculele sinaptice în fanta sinaptică. Veziculele sinaptice fuzionează cu membrana presinaptică în timpul acestui proces de exocitoză. Membranele veziculelor vechi devin parte a membranei presinaptice, iar noile vezicule se desprind dintr-o zonă adiacentă a membranei. Aceste vezicule noi sunt ulterior reumplute cu neurotransmițători nou-sintetizați sau „reciclați” .
Neurotransmițătorii eliberați difuzează prin fanta sinaptică îngustă. La nivelul membranei postsinaptice, moleculele de neurotransmițător se leagă de moleculele receptoare legate de membrană cu situsuri de recunoaștere specifice pentru acel neurotransmițător. Legarea neurotransmițătorului de receptor declanșează un răspuns postsinaptic specific pentru acel receptor. Aceste răspunsuri pot fi fie excitatorii, fie inhibitorii, în funcție de proprietățile receptorului. Dacă stimularea receptorului are ca rezultat faptul că membrana postsinaptică devine mai pozitivă din punct de vedere electric (depolarizată), este vorba de un potențial postsinaptic excitator (EPSP). Dacă este mai negativ (hiperpolarizat), acesta este un potențial postsinaptic inhibitor (IPSP). Excitarea și inhibiția depind de proprietățile receptorului și nu de neurotransmițător. Receptorii cuplați la canalele de sodiu sau de calciu sunt excitanți și produc o depolarizare a membranei postsinaptice, în timp ce receptorii cuplați la canalele de clorură sau de potasiu sunt inhibitori și produc o hiperpolarizare a membranei postsinaptice. Astfel de receptori cuplați la canale ionice se numesc receptori ionotropi.
Alți receptori sunt cuplați la sisteme de „al doilea mesager” care inițiază o serie de reacții biochimice în celula postsinaptică. Aceștia sunt receptorii metabotropi . Receptorii metabotropici pot produce multe
evenimente postsinaptice. Acestea variază de la activarea directă a canalelor ionice adiacente, la alterarea sensibilității receptorilor, la transcrierea unor acizi ribonucleici mesageriali (ARN) specifici sau chiar activarea unor gene specifice . Sinapsele chimice fac parte dintr-un sistem de comunicare foarte adaptabil și flexibil. Acestea nu sunt structuri anatomice statice cu proprietăți fixe, ci sunt structuri dinamice, capabile să își modifice proprietățile moleculare în funcție de schimbarea circumstanțelor.
Există literalmente sute de neurotransmițători. Unii sunt compuși destul de simpli, cum ar fi acetilcolina, serotonina, catecolaminele (dopamina, norepinefrina și epinefrina) și o serie de aminoacizi . Mulți sunt mai complecși și fac parte din vasta gamă de transmițători neuropeptidici. Odată eliberați în fanta sinaptică, neurotransmițătorii rămân activi până când sunt fie modificați chimic, fie preluați înapoi în butonul sinaptic de către sisteme speciale de transport și reciclați. La sinapsele colinergice, acetilcolinesteraza este prezentă în fanta sinaptică. Această enzimă scindează neurotransmițătorul în acetat și colină, dintre care niciunul nu este activ. Serotonina și epinefrina, pe de altă parte, sunt preluate în terminalul presinaptic și reciclate.
Lasă un răspuns