Subphylum Crustacea
Natura generală a unui crustaceu
Crustaceele diferă de alte artropode într-o varietate de moduri, dar singura caracteristică cu adevărat distinctivă este că crustaceele sunt singurele artropode cu două perechi de antene. În plus față de cele două perechi de antene și o pereche de mandibule, crustaceele au două perechi de maxilare pe cap, urmate de o pereche de apendice pe fiecare segment de corp sau somit. La unele crustacee, nu toate somitele poartă apendice. Toate apendicele, cu excepția, probabil, a primelor antene, sunt primitiv biramoase (două ramuri principale), iar cel puțin unele apendice ale adulților actuali prezintă această condiție. Organele specializate pentru respirație, dacă sunt prezente, funcționează ca branhii.
Majoritatea crustaceelor au între 16 și 20 de somite, dar unele forme au 60 de somite sau mai mult. Un număr mai mare de somite este o caracteristică primitivă. Condiția mai derivată este aceea de a avea mai puține segmente și o tagmație crescută. Tagmatele majore sunt capul, toracele și abdomenul, dar acestea nu sunt omoloage în întreaga clasă (sau chiar în cadrul unor subclase) din cauza gradului variabil de fuziune a somitelor, de exemplu, ca în cazul cefalotorace.
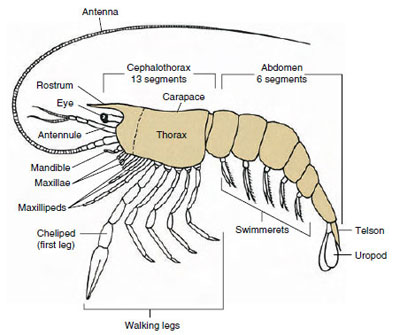
Figura 19-1
Planul arhetipal al Malacostraca. Cele două maxilare și cele trei
maxilipede au fost separate schematic pentru a ilustra planul
general.
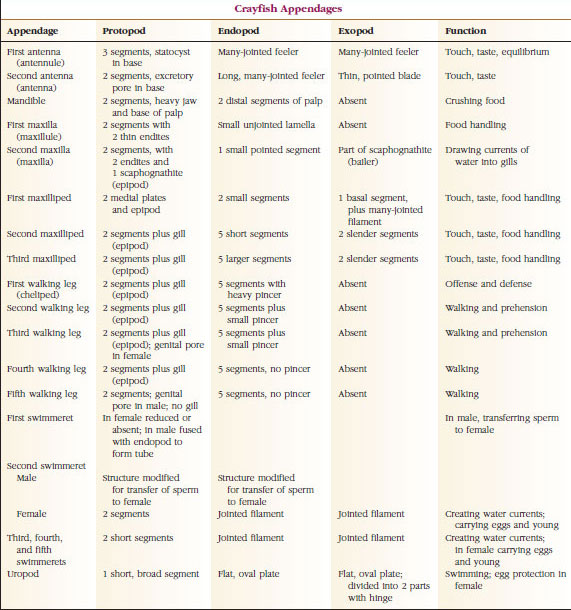
De departe, cel mai mare grup de crustacee este clasa Malacostraca, care include homari, crabi, creveți, săritori de plajă, scroafe și multe altele. Aceștia prezintă un aranjament surprinzător de constant al segmentelor corpului și al tagmei, care este considerat planul ancestral al clasei (figura 19-1). Acest plan corporal tipic are un cap format din cinci somite fuzionate (șase la nivel embrionar), un torace de opt somite și un abdomen de șase somite (șapte la câteva specii). La capătul anterior se află un rostru nesegmentat, iar la capătul posterior se află un telson nesegmentat, care împreună cu ultimul somit abdominal și cu uropodele sale formează un evantai al cozii în multe forme.
La multe crustacee, cuticula dorsală a capului se poate extinde posterior și în jurul laturilor animalului pentru a acoperi sau a fi fuzionată cu o parte sau cu toate somitele toracice și abdominale. Acest înveliș se numește carapace. La unele grupuri, carapacea formează valve asemănătoare unor scoici care acoperă cea mai mare parte sau întregul corp. La decapode (inclusiv homari, creveți, crabi și altele), carapacea acoperă întregul cefalotorace, dar nu și abdomenul.
Formă și funcție
Din cauza mărimii lor și a disponibilității facile, crustaceele mari, cum ar fi racii, au fost studiate mai mult decât alte grupuri. Ele sunt, de asemenea, frecvent studiate în cadrul cursurilor introductive de laborator. Prin urmare, multe dintre comentariile care urmează se aplică în mod specific racilor și rudelor lor.
Caracteristici externe
Corpurile crustaceelor sunt acoperite cu o cuticulă secretată, compusă din chitină, proteine și material calcaros. Plăcile mai dure și grele ale crustaceelor mai mari sunt deosebit de bogate în depozite calcaroase. Învelișul protector dur este moale și subțire la nivelul articulațiilor dintre somite, permițând flexibilitatea mișcărilor. Carapacea, dacă este prezentă, acoperă o mare parte sau întregul cefalotorace; la decapode, cum ar fi racii, toate segmentele capului și toracice sunt închise dorsal de carapace. Fiecare somit care nu este înconjurat de carapace este acoperit de o placă cuticulară dorsală, sau tergum (figura 19-2A), iar o bară transversală ventrală, sternul, se află între apendicele segmentare (figura 19-2B). Abdomenul se termină cu un telson, care nu este considerat un somit și care poartă anusul. (Este posibil ca telsonul să fie omologul pygidium-ului anelidelor.)
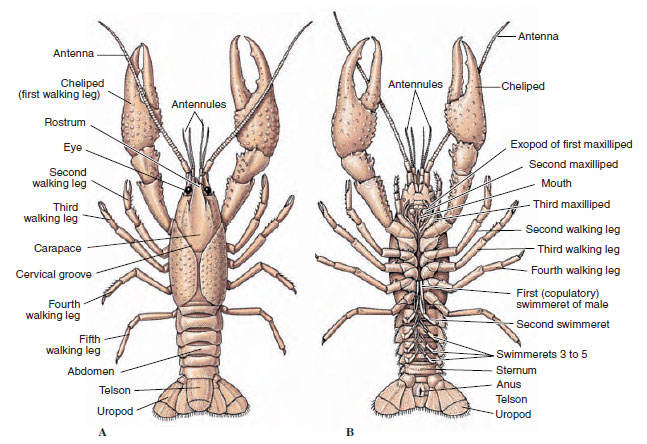
Figura 19-2
Structura externă a racilor. A, vedere dorsală. B, vedere ventrală.
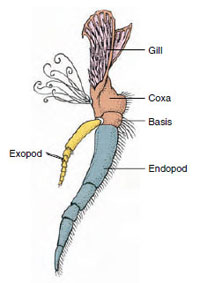
Figura 19-3
Părți ale unui
apendice de crustaceu biramat (al treilea maxiliped al unui
crayfish).
Poziția gonoporilor variază în funcție de sex și de grupul de crustacee. Ei pot fi pe sau la baza unei perechi de apendice, la capătul terminal al corpului sau pe somite fără picioare. La raci, deschiderile vaselor deferente se află pe partea mediană, la baza celei de-a cincea perechi de picioare umblătoare, iar cele ale oviductului se află la baza celei de-a treia perechi. La femele, orificiul receptaculului seminal este situat, de obicei, pe linia medianăventrală, între a patra și a cincea pereche de picioare de mers.
Apendice: Membrii claselor Malacostraca (inclusiv racii) și Remipedia au de obicei o pereche de apendice articulate pe fiecare somit (figura 19-3), deși somitele abdominale din celelalte clase nu poartă apendice. O specializare considerabilă este evidentă în cazul apendicelor crustaceelor derivate, cum ar fi racii. Cu toate acestea, toate sunt variații ale planului de bază, biramous, ilustrat de un apendice al racilor, cum ar fi maxilipedul (un membru toracic modificat pentru a deveni un apendice al capului) (figurile 19-3 și 19-4). Porțiunea bazală, sau protopodul, poartă un exopod lateral și un endopod medial. Protopodul este alcătuit din una sau două articulații (baza și coxa), în timp ce exopodul și endopodul au de la una până la mai multe articulații fiecare. Unele apendice, cum ar fi picioarele de mers ale racilor, au devenit secundar unirame. Pe membrele crustaceelor apar uneori procese mediale sau laterale, numite endite și, respectiv, exite, iar un exite pe protopod se numește epipod. Epipodele sunt adesea modificate ca branhii. Tabelul 19-1 prezintă modul în care diversele apendice s-au modificat față de planul biramat pentru a corespunde unor funcții specifice.
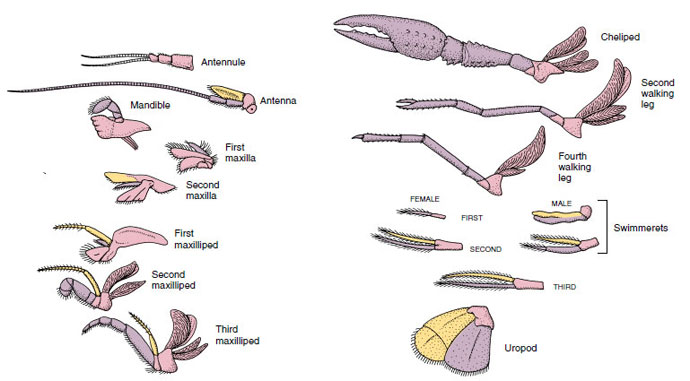
Figura 19-4
Anexele unui rac arătând modul în care au devenit modificate față de planul biramat de bază, așa cum se găsește la un
swimmeret. Protopodul, roz; endopodul, violet; exopodul, galben.
Structurile care au un plan de bază asemănător și care au descins dintr-o formă comună se spune că sunt omoloage, indiferent dacă au aceeași funcție sau nu. Având în vedere că picioarele specializate pentru mers, aparatele bucale, chelipedii și mrejele de înot s-au dezvoltat toate dintr-un tip comun de biramă, dar s-au modificat pentru a îndeplini funcții diferite, toate sunt omoloage între ele, o condiție cunoscută sub numele de omologie serială. În mod primitiv, membrele erau toate foarte asemănătoare, dar în timpul evoluției modificărilor structurale, unele ramuri au fost reduse, unele au dispărut, altele au fost mult modificate, iar unele părți noi au fost adăugate. Racii și aliații lor posedă cea mai elaborată omologie serială din regnul animal, având 17 tipuri de apendice distincte, dar omoloage în serie (tabelul 19-1).
TABELUL 19.1
Caracteristici interne
Sistemele muscular și nervos și segmentarea în torace și abdomen arată în mod clar un metamerism moștenit de la strămoșii de tip anelide, dar există modificări marcante în alte sisteme. Cele mai multe modificări implică concentrarea unor părți într-o anumită regiune sau altfel reducerea sau pierderea completă a unor părți, cum ar fi intersepta.
Hemocoel: Spațiul corporal major la artropode nu este celomul, ci un hemocoel plin de sânge. În timpul dezvoltării embrionare a majorității artropodelor, cavități celomice vestigiale se deschid în mezodermul a cel puțin unor somite. Acestea sunt în curând șterse sau devin continue cu spațiul dintre structurile mezodermice și ectodermice în curs de dezvoltare și gălbenuș. Acest spațiu devine hemocoel și, prin urmare, nu este acoperit de un peritoneu mezodermal. La crustacee, singurele compartimente celomice care rămân sunt sacii terminali ai organelor excretoare și spațiul din jurul gonadelor.
Sistemul muscular: Mușchii striați alcătuiesc o parte considerabilă a corpului majorității crustaceelor. Mușchii sunt de obicei dispuși în grupuri antagoniste: flexori, care atrag o parte spre corp, și extensori, care o extind spre exterior. Abdomenul unui rac are flexori puternici (figura 19-5), care sunt folosiți atunci când animalul înoată în spate – cel mai bun mijloc de scăpare al său. Mușchii puternici de o parte și de alta a stomacului controlează mandibulele.
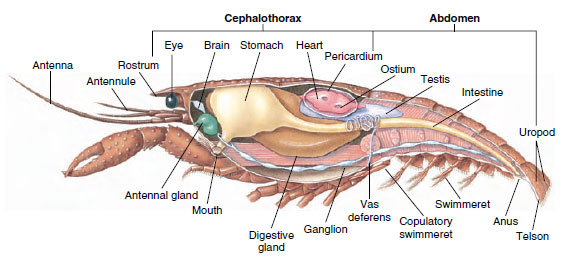
Figura 19-5
Structura internă a unui rac mascul.
Sistemul respirator: Schimbul de gaze respiratorii la crustaceele mai mici are loc pe zone mai subțiri de cuticulă (de exemplu, în picioare) sau pe întregul corp, iar structurile specializate pot fi absente. Crustaceele mai mari au branhii, care sunt proiecții delicate, asemănătoare unor pene, cu o cuticulă foarte subțire. La decapode, părțile laterale ale carapacei înconjoară cavitatea branhială, care este deschisă anterior și ventral (figura 19-6). Branhiile se pot proiecta din peretele pleural în cavitatea branhială, de la articulația picioarelor toracice cu corpul sau de la coxele toracice. Ultimele două tipuri sunt tipice pentru raci. „Bălăciorul”, o parte a celui de-al doilea maxilar, atrage apa peste filamentele branhiale, în cavitatea branhială la bazele picioarelor și o scoate din cavitatea branhială la partea anterioară.
Sistemul circulator: Crustaceele și alte artropode au un sistem circulator de tip „deschis” sau lacunar. Acest lucru înseamnă că nu există vene și nu există o separare a sângelui de lichidul interstițial, așa cum se întâmplă la animalele cu sisteme închise. Hemolimfa (sângele) părăsește inima prin intermediul arterelor, circulă prin hemocoel și revine în sinusurile venoase, sau spațiile, în loc de vene, înainte de a reintra în inimă. Annelidele au un sistem închis, la fel ca și vertebratele.
O inimă dorsală este principalul organ de propulsie. Este un sac cu o singură cameră de mușchi striat. Hemolimfa intră în inimă din sinusul pericardic înconjurător prin ostii perechi, cu supape care împiedică refluxul în sinus (figura 19-6). Din inimă, hemolimfa intră în una sau mai multe artere. Supapele din artere împiedică un reflux al hemolimfei. Arterele mici se varsă în sinusurile tisulare, care, la rândul lor, se descarcă adesea într-un sinus sternal mare (figura 19-6).
De acolo, canalele aferente sinusurilor transportă hemolimfa către branhii, dacă sunt prezente, pentru schimbul de oxigen și dioxid de carbon. Hemolimfa se întoarce apoi în sinusul pericardic prin canale eferente (figura 19-6).
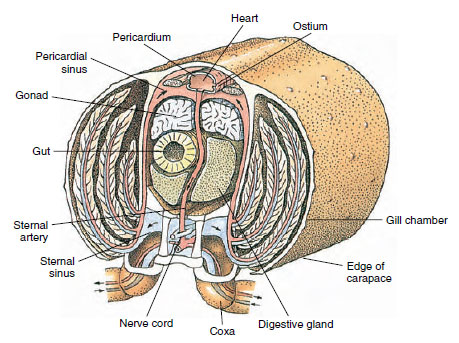
Figura 19-6
Secțiune transversală diacritică prin regiunea cardiacă a unui rac arătând direcția fluxului sanguin în acest sistem
sângeros „deschis”
. Inima pompează sângele către țesuturile corpului prin artere, care se varsă în sinusurile tisulare. Sângele
sângele care se întoarce intră în sinusul sternal, apoi trece prin branhii pentru schimbul de gaze și, în final, se întoarce în sinusul pericardic prin canale
eferente. Observați absența venelor.
Hemolimfa la artropode este în mare parte incoloră. Ea include celule ameboide de cel puțin două tipuri. Hemocianina, un pigment respirator care conține cupru, sau hemoglobina, un pigment care conține fier, pot fi transportate în soluție. Hemolimfa are proprietatea de a se coagula, ceea ce împiedică pierderea ei în cazul unor leziuni minore. Unele celule ameboide eliberează un coagulant asemănător trombinei, care precipită coagularea.
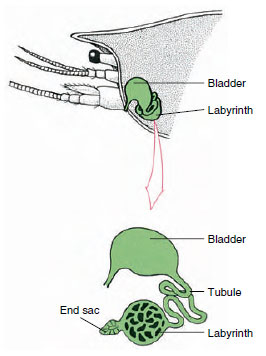
Figura 19-7
Schema glandei antenale (glanda verde) a
racilor. (În poziție naturală organul este mult
pliat.) Unele crustacee nu au labirint,
și tubul excretor (canalul nefridial) este
un tub mult încolăcit.
Sistemul excretor: Organele excretoare ale crustaceelor adulte sunt o pereche de structuri tubulare situate în partea ventrală a capului, anterior esofagului (figura 19-5). Ele se numesc glande antenale sau glande maxilare, în funcție de faptul că se deschid la baza antenelor sau a celui de-al doilea maxilar. Câteva crustacee adulte le au pe amândouă. Organele excretoare ale decapodelelor sunt glandele antenale, numite și glande verzi în acest grup. Crustaceele nu au tubuli malpighieni, organele excretoare ale păianjenilor și insectelor.
Sacul terminal al glandei antennale, care provine dintr-un compartiment celomic embrionar, este format dintr-o veziculă mică (saccule) și o masă spongioasă numită labirint. Labirintul se conectează printr-un tubul excretor la o vezică dorsală, care se deschide spre exterior printr-un por situat pe suprafața ventrală a segmentului antennal bazal (figura 19-7). Presiunea hidrostatică din interiorul hemocoelului asigură forța de filtrare a lichidului în sacul terminal. Pe măsură ce filtratul trece prin tubul excretor și vezica urinară, este modificat prin resorbția de săruri, aminoacizi, glucoză și o parte din apă și este în final excretat sub formă de urină.
Excreția deșeurilor azotate (în principal amoniac) are loc prin difuzie prin zone subțiri de cuticulă, în special branhiile, iar așa-numitele organe excretoare funcționează în principal pentru a regla compoziția ionică și osmotică a fluidelor corporale. Crustaceele de apă dulce, cum ar fi racii de mare, sunt în mod constant amenințate de supradiluarea sângelui lor cu apă, care difuzează prin branhii și alte suprafețe permeabile la apă. Glandele verzi, prin formarea unei urini diluate, cu conținut scăzut de sare, acționează ca un dispozitiv eficient de „control al inundațiilor”. O parte din Na+ și Cl- se pierd în urină, dar această pierdere este compensată prin absorbția activă a sării dizolvate de către branhii. La crustaceele marine, cum ar fi homarii și crabii, rinichii au rolul de a regla compoziția salină a hemolimfei prin modificarea selectivă a conținutului de sare din urina tubulară. În aceste forme, urina rămâne izosmotică față de sânge.
Sistemele nervos și senzorial: Sistemele nervoase ale crustaceelor și ale anelidelor au multe în comun, deși cele ale crustaceelor au mai multe fuziuni de ganglioni (figura 19-5). Creierul este o pereche de ganglioni supraesofagieni care furnizează nervii către ochi și două perechi de antene. El este unit prin conectoare cu ganglionul subesofagian, o fuziune a cel puțin cinci perechi de ganglioni care furnizează nervi către gură, anexe, esofag și glande antenale. Cordonul nervos ventral dublu are o pereche de ganglioni pentru fiecare somit și nervi care deservesc apendicele, mușchii și alte părți.
În plus față de acest sistem central, poate exista un sistem nervos simpatic asociat cu tractul digestiv.
Crustaceele au organe de simț mai bine dezvoltate decât aneliții. Cele mai mari organe de simț ale racilor sunt ochii și statocitele. Distribuite pe scară largă pe corp sunt firele de păr tactil, proiecții delicate de cuticulă care sunt deosebit de abundente pe chelae, aparatele bucale și telson. Simțurile chimice ale gustului și mirosului se regăsesc în firele de păr de pe antene, aparatele bucale și din alte locuri.
Un statocist asemănător unui sac, care se deschide la suprafață printr-un por dorsal, se găsește pe segmentul bazal al fiecărei prime antene a racilor de mare. Statocistul conține o creastă care poartă peri senzoriali formați din căptușeala chitinoasă și boabe de nisip care servesc drept statoliți. Ori de câte ori animalul își schimbă poziția, modificările corespunzătoare ale poziției granulelor de pe firele de păr senzoriale sunt transmise ca stimuli către creier, iar animalul se poate adapta în consecință. Fiecare mutare (ecdysis) a cuticulei duce la pierderea căptușelii cuticulare a statocistului și, odată cu ea, a granulelor de nisip. Noile granule sunt preluate prin porul dorsal după ecdiză.
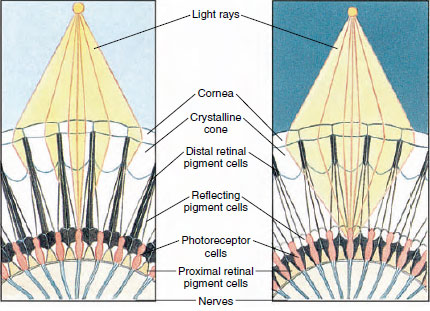
Figura 19-8
Porțiune din ochiul compus al unui artropod care arată migrarea pigmentului
în ommatidii pentru vederea diurnă și nocturnă. Cinci ommatidii sunt reprezentate în
fiecare diagramă. În timpul zilei, fiecare ommatidiu este înconjurat de un colier
pigmentar întunecat, astfel încât fiecare ommatidiu este stimulat doar de razele de lumină
care intră în propria cornee (viziune mozaicată); în timpul nopții, pigmentul formează coliere
incomplete și razele de lumină se pot răspândi la ommatidii adiacente
(imagine continuă, sau de suprapunere).
Ochii la multe crustacee sunt compuși, compuși din mai multe unități fotoreceptoare numite ommatii (figura 19-8). Acoperind suprafața rotunjită a fiecărui ochi se află o zonă transparentă de cuticulă, corneea, care este împărțită în multe pătrate mici sau hexagoane cunoscute sub numele de fațete. Aceste fațete reprezintă capetele exterioare ale ommatidiilor. Fiecare ommatidiu se comportă ca un ochi mic și conține mai multe tipuri de celule dispuse în formă de coloană (figura 19-8). Între ommatidiile adiacente se găsesc celule pigmentare negre.
Mișcarea pigmentului într-un ochi compus al artropodelor îi permite acestuia să se adapteze la diferite cantități de lumină. Există trei seturi de celule pigmentare în fiecare ommatidiu: retinale distale, retinale proximale și reflectante; acestea sunt dispuse astfel încât pot forma un guler sau un manșon mai mult sau mai puțin complet în jurul fiecărui ommatidiu. În cazul unei adaptări puternice la lumină sau la zi, pigmentul retinian distal se deplasează spre interior și se întâlnește cu pigmentul retinian proximal care se deplasează spre exterior, astfel încât se formează un manșon pigmentar complet în jurul ommatidiului (figura 19-8). În această stare, numai razele care lovesc direct corneea vor ajunge la celulele fotoreceptoare (retinulare), deoarece fiecare ommatidiu este protejat de celelalte. Astfel, fiecare ommatidiu va vedea doar o zonă limitată a câmpului vizual (o imagine mozaicată sau de apoziție). În lumină slabă, pigmenții distali și proximali se separă, astfel încât razele de lumină, cu ajutorul celulelor pigmentare reflectante, au șansa de a se răspândi la ommatidiile adiacente și de a forma o imagine continuă, sau de suprapunere. Acest al doilea tip de viziune este mai puțin precis, dar profită la maximum de cantitatea limitată de lumină primită.
Reproducere, cicluri de viață și funcție endocrină
Majoritatea crustaceelor au sexe separate și există diverse specializări pentru copulație în cadrul diferitelor grupuri. Cârtițele sunt monoice, dar în general practică fertilizarea încrucișată. La unele ostracode masculii sunt rari, iar reproducerea este de obicei partenogenetică. Majoritatea crustaceelor își clocesc ouăle într-un anumit mod: branchiopodele și cracii au camere de clocit speciale, copepodele au saci de clocit atașați de părțile laterale ale abdomenului (a se vedea figura 19-19), iar mulți malacostracși poartă ouăle și puii atașați de apendicele abdominale.
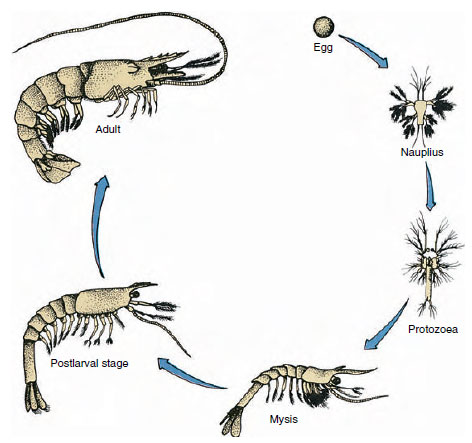
Figura 19-9
Ciclul de viață al unui crevete de golf Penaeus. Penaeidele își depun icrele la adâncimi cuprinse între 40 și 90 m. Formele larvare tinere sunt
planctonice și se deplasează spre țărm, în ape cu salinitate mai scăzută, pentru a se dezvolta ca puiet. Creveții mai bătrâni se întorc în apă mai adâncă
în larg.
Craioții au o dezvoltare directă: nu există forme larvare. Un micuț puiet cu aceeași formă ca și adultul și cu un set complet de apendice și somite eclozează din ou. Cu toate acestea, dezvoltarea este indirectă la majoritatea crustaceelor, iar din ou eclozează o larvă destul de diferită de cea a adultului ca structură și aspect. Trecerea de la larvă la adult se numește metamorfoză. Larva primitivă și cea mai răspândită la crustacee este naupliul (figurile 19-9 și 19-23). Nauplii poartă doar trei perechi de apendice: primele antene unirame, antenele birame și mandibulele birame. Toate funcționează ca apendice de înot în acest stadiu.
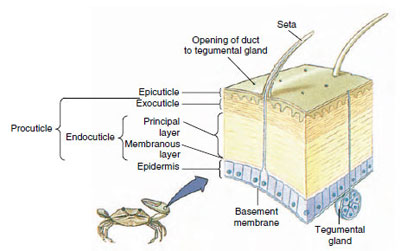
Figura 19-10
Structura cuticulei crustaceelor.
Dezvoltarea ulterioară poate implica o schimbare treptată a formei corporale adulte, iar apendicele și somitele sunt adăugate printr-o serie de moli, sau asumarea formei adulte poate implica schimbări mai abrupte. De exemplu, metamorfoza unui barnacle trece de la un nauplius care înoată liber la o larvă cu o carapace bivalvă numită ciprid și, în cele din urmă, la un adult sesil cu plăci calcaroase.
Ecdiza: Ecdiza (ek´duh-sis) (Gr. ekdyein, a se dezbrăca), sau mutația, este necesară pentru ca organismul să crească în dimensiune deoarece exoscheletul nu este viu și nu crește odată cu creșterea animalului. O mare parte din funcționarea unui crustaceu, inclusiv reproducerea, comportamentul și multe procese metabolice, este direct afectată de fiziologia ciclului de mută.
Cuticul, care este secretat de epiderma subiacentă, are mai multe straturi (figura 19-10). Cel mai exterior este epicuticula, un strat foarte subțire de proteină impregnată de lipide. Cea mai mare parte a cuticulei este reprezentată de cele câteva straturi de procuticulă: (1) exocuticulă, care se află chiar sub epicuticulă și conține proteine, săruri de calciu și chitină; (2) endocuticulă, care la rândul ei este compusă din (3) un strat principal, care conține mai multă chitină și mai puține proteine și este puternic calcificat, și (4) un strat membranos necalcificat, un strat relativ subțire de chitină și proteine.
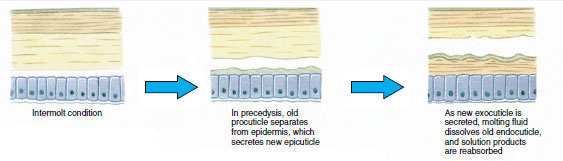
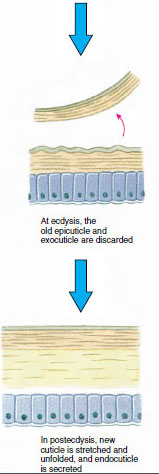
Cu ceva timp înainte de ecdiza propriu-zisă, celulele epidermice se măresc considerabil. Ele se separă de stratul membranos, secretă o nouă epicuticulă și încep să secrete o nouă exocuticulă (figura 19-11). Enzimele sunt eliberate în zona de deasupra noii epicuticle. Aceste enzime încep să dizolve vechea endocuticulă, iar produsele solubile sunt resorbite și depozitate în interiorul corpului crustaceului. Unele săruri de calciu sunt stocate sub formă de gastrolite (acumulări minerale) în pereții stomacului. În cele din urmă, din vechea cuticulă rămân doar exocuticula și epicuticula, sub care se află noua epicuticulă și noua exocuticulă. Animalul înghite apă, pe care o absoarbe prin intestin, iar volumul de sânge crește foarte mult. Presiunea internă face ca cuticula să se despice, iar animalul se scoate singur din vechiul său exoschelet (figura 19-12). Urmează apoi o întindere a noii cuticule încă moi, depunerea noii endocuticule, redepunerea sărurilor anorganice recuperate și a altor constituenți și întărirea noii cuticule. În timpul perioadei de mută, animalul este lipsit de apărare și rămâne ascuns.
 |
|
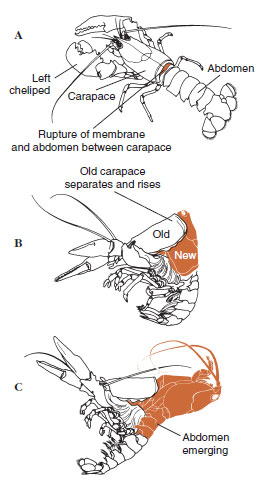 |
 |
| Figura 19-12 Secvența de muiere la homar, Homarus americanus. A, Membrana dintre carapace și abdomen se rupe, iar carapacea începe să se ridice lent. Această etapă poate dura până la 2 ore. B și C, Capul, toracele și, în cele din urmă, abdomenul se retrag. Acest proces durează de obicei nu durează mai mult de 15 minute. Imediat după ecdiză, cheliepedele sunt desecate și corpul este foarte moale. Homarul continuă absorbția rapidă de apă, astfel încât în 12 ore corpul crește cu aproximativ 20% în lungime și 50% în greutate. Apa din țesuturi va fi înlocuită de proteine în săptămânile următoare. |
Figura 19-11 Secreția și resorbția de gușă în ecdiză. |
Când un crustaceu este tânăr, ecdiza trebuie să aibă loc frecvent pentru a permite creșterea, iar ciclul de muiere este relativ scurt. Pe măsură ce animalul se apropie de maturitate, perioadele de intermoltare devin progresiv mai lungi, iar la unele specii mutul încetează cu totul. În timpul perioadelor de intermitență, are loc o creștere a masei tisulare pe măsură ce țesutul viu înlocuiește apa.
Controlul hormonal al ciclului de ecdiză
Deși ecdiza este controlată hormonal, ciclul este adesea inițiat de un stimul de mediu perceput de sistemul nervos central. Astfel de stimuli pot include temperatura, lungimea zilei și umiditatea (în cazul crabilor de uscat). Semnalul de la sistemul nervos central diminuează producția unui hormon de inhibare a mucegaiului de către organul X. Organul X este un grup de celule neurosecretoare din medulla terminalis a creierului. La raci și la alte decapode, medulla terminalis se găsește în pedunculul ocular. Hormonul este transportat în axonii organului X până la glanda sinus (care, probabil, nu are ea însăși o funcție glandulară), aflată tot în pedunculul ocular, unde este eliberat în hemolimfă.
O scădere a nivelului hormonului inhibitor de mucegai favorizează eliberarea unui hormon de mucegai din organul Y. Organele Y se află sub epidermă, în apropierea mușchilor adductori ai mandibulei, și sunt omoloage cu glandele protoracice ale insectelor, care produc hormonul ecdysone. Acțiunea hormonului de mută este de a iniția procesele care duc la ecdysis (proecdysis). Odată inițiat, ciclul se desfășoară automat, fără acțiunea ulterioară a hormonilor din partea organelor X sau Y.
Alte funcții endocrine: Nu numai că îndepărtarea eyalelor accelerează muierea, dar s-a constatat, de asemenea, cu peste 100 de ani în urmă, că crustaceele ale căror eyalelor au fost îndepărtate nu mai pot ajusta colorația corpului la condițiile de fond. Cu peste 50 de ani în urmă, s-a descoperit că acest defect nu este cauzat de pierderea vederii, ci de pierderea hormonilor din ehipelaguri. Culoarea corpului crustaceelor este în mare parte rezultatul pigmenților din celulele speciale ramificate (cromatofori) din epidermă.
Concentrarea granulelor de pigment în centrul celulelor determină un efect de iluminare, iar dispersarea pigmentului în toate celulele determină un efect de întunecare. Comportamentul pigmentului este controlat de hormoni din celulele neurosecretoare din pedunculul ocular, la fel ca și migrarea pigmentului retinian pentru adaptarea la lumină și la întuneric în ochi (figura 19-8).
Liberarea de material neurosecretor din organele pericardice din peretele pericardului determină o creștere a ritmului și amplitudinii bătăilor inimii.
Glandele androgenice, descoperite pentru prima dată la un amfipod (Orchestia, un țopârlan comun de pe plajă), apar la masculii malacostracieni. Spre deosebire de majoritatea celorlalte organe endocrine de la crustacee, acestea nu sunt organe neurosecretoare. Secreția lor stimulează exprimarea caracteristicilor sexuale masculine. Tinerii malacostracani au glande androgenice rudimentare, dar la femele aceste glande nu se dezvoltă. Dacă acestea sunt implantate în mod artificial la o femelă, ovarele acesteia se transformă în testicule și încep să producă spermă, iar apendicele sale încep să capete caracteristici masculine la următoarea mutare. La izopode, glandele androgenice se găsesc în testicule; la toți ceilalți malacostracani, ele se află între mușchii coxopodelor ultimelor picioare toracice și sunt parțial atașate lângă capetele vaselor deferente. Deși femelele nu posedă organe similare glandelor androgenice, ovarele lor produc unul sau doi hormoni care influențează caracteristicile sexuale secundare.
Este posibil să fie prezenți hormoni care influențează alte procese corporale la crustacee, iar dovezile sugerează că o substanță neurosecretoare produsă în pedunculul ocular reglează nivelul zahărului din sânge.
Habitudini de hrănire
Habitudinile de hrănire și adaptările pentru hrănire variază foarte mult în rândul crustaceelor. Multe forme pot trece de la un tip de hrănire la altul în funcție de mediu și de disponibilitatea hranei, dar toate folosesc același set fundamental de organe bucale. Mandibulele și maxilarele au rolul de a ingera hrana; maxilipedele țin și zdrobesc hrana. La prădători, picioarele umblătoare, în special chelipedele, servesc la capturarea hranei.
Multe crustacee, atât mari cât și mici, sunt prădătoare, iar unele au adaptări interesante pentru uciderea prăzii. O formă asemănătoare creveților, Lygiosquilla, are pe unul dintre picioarele de mers un deget specializat care poate fi tras într-o canelură și eliberat brusc pentru a străpunge prada care trece. Creveții pistol (Alpheus spp.) au o chela enorm de mărită care poate fi armată ca ciocanul unui pistol și pocită cu o forță care le amețește prada.
Hrană pentru cei care se hrănesc în suspensie variază de la plancton și detritus până la bacterii. Prădătorii consumă larve, viermi, crustacee, melci și pești. Scafandrii mănâncă materie animală și vegetală moartă. Cei care se hrănesc în suspensie, cum ar fi creveții zână, puricii de apă și cracii, își folosesc picioarele, care poartă o franjură groasă de setae, pentru a crea curenți de apă care mătură particulele de hrană prin setae. Creveții de noroi (Upogebia spp.) folosesc setae lungi pe primele două perechi de apendice toracice pentru a strecura materialul alimentar din apa care circulă prin vizuina lor prin mișcări ale mreterelor de înot.
Creveții au un stomac în două părți (figura 19-13). Prima parte conține o moară gastrică în care hrana, deja sfâșiată de mandibule, poate fi măcinată în continuare de trei dinți calcaroși în particule suficient de fine pentru a trece printr-un filtru setos din a doua parte; particulele de hrană trec apoi în intestin pentru digestia chimică.
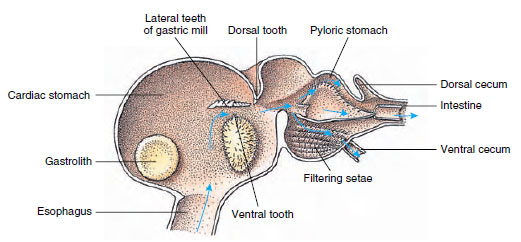
Figura 19-13
Stomacul Malacostracanului care arată „moara” gastrică și direcțiile de deplasare a alimentelor. Moara are creste chitinoase, sau
dinți, pentru masticație, și setae pentru strecurarea alimentelor înainte de a trece în stomacul piloric.
.
Lasă un răspuns