Subphylum Crustacea
General Nature of a Crustacea
Skorupiaki różnią się od innych stawonogów na wiele sposobów, ale jedyną naprawdę wyróżniającą cechą jest to, że skorupiaki są jedynymi stawonogami z dwiema parami czułków. Oprócz dwóch par czułków i pary żuchw, skorupiaki mają dwie pary szczęk na głowie, a następnie parę przydatków na każdym segmencie ciała lub somitu. U niektórych skorupiaków nie wszystkie somity mają wyrostki. Wszystkie wyrostki, może z wyjątkiem pierwszych czułków, są pierwotnie dwurożne (dwie główne gałęzie), a przynajmniej niektóre wyrostki współczesnych osobników dorosłych wykazują ten stan. Organy wyspecjalizowane do oddychania, jeśli są obecne, funkcjonują jako skrzela.
Większość skorupiaków ma od 16 do 20 somitów, ale niektóre formy mają 60 somitów lub więcej. Większa liczba somitów jest cechą prymitywną. Bardziej pochodnym warunkiem jest posiadanie mniejszej liczby segmentów i zwiększonej tagmatyzacji. Główne tagmaty to głowa, tułów i odwłok, ale nie są one homologiczne w całej klasie (a nawet w niektórych podklasach) ze względu na różny stopień zrastania się somitów, na przykład jak u głowotułowia.
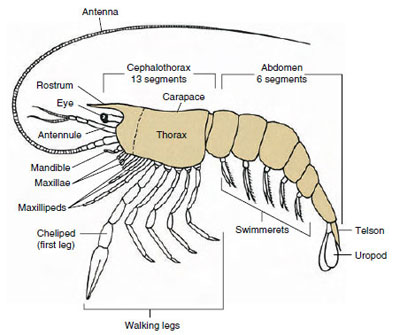
Ryc. 19-1
Archetypowy plan Malacostraca. Dwa szczęki i trzy
maksylitopsy zostały rozdzielone schematycznie, aby zilustrować
plan ogólny.
Zdecydowanie największą grupą skorupiaków jest klasa Malacostraca, do której należą homary, kraby, krewetki, lejkowce plażowe, siewkowce i wiele innych. Wykazują one zaskakująco stały układ segmentów ciała i tagmatów, który jest uważany za plan przodków tej klasy (rysunek 19-1). Ten typowy plan budowy ciała składa się z głowy złożonej z pięciu (sześciu embrionalnie) połączonych somitów, tułowia z ośmiu somitów i odwłoka z sześciu somitów (u kilku gatunków z siedmiu). Na przednim końcu znajduje się niesegmentowane rostrum, a na tylnym niesegmentowany telson, który z ostatnim somitem brzusznym i jego uropodiami tworzy w wielu formach wachlarz ogonowy.
W wielu skorupiakach kutikula grzbietowa głowy może rozciągać się ku tyłowi i wokół boków zwierzęcia, pokrywając lub łącząc się z niektórymi lub wszystkimi somitami piersiowymi i brzusznymi. Pokrycie to nazywane jest pancerzem. U niektórych grup pancerzyki tworzą zawory przypominające muszle małży, które pokrywają większość lub całość ciała. W decapoda (w tym homary, krewetki, kraby i inne), pancerz obejmuje cały głowotułów, ale nie odwłok.
Forma i funkcja
Z powodu ich wielkości i łatwej dostępności, duże skorupiaki, takie jak raki, były badane bardziej niż inne grupy. Są one również powszechnie badane w ramach wstępnych kursów laboratoryjnych. Dlatego wiele z poniższych uwag odnosi się w szczególności do raków i ich krewnych.
Cechy zewnętrzne
Ciała skorupiaków pokryte są wydzielaną kutykulą składającą się z chityny, białka i materiału wapiennego. Twardsze, ciężkie płyty większych skorupiaków są szczególnie bogate w osady wapienne. Twarda powłoka ochronna jest miękka i cienka w miejscach połączeń między somitami, co umożliwia elastyczność ruchów. Pancerz, jeśli występuje, okrywa większą część lub całość głowotułowia; u dziesięciornic, takich jak raki, wszystkie segmenty głowowe i piersiowe są zamknięte grzbietowo przez pancerz. Każdy somit nieobjęty pancerzem pokryty jest grzbietową płytką kutikularną lub tergum (rysunek 19-2A), a pomiędzy wyrostkami segmentalnymi znajduje się brzuszna poprzeczna listwa, mostek (rysunek 19-2B). Odwłok kończy się telsonem, który nie jest uważany za somit i zawiera odbyt. (Telson może być homologiczny do pygidium annelidów)
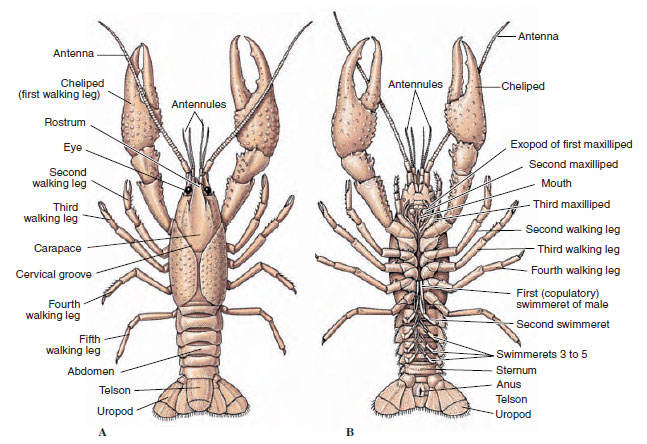
Rysunek 19-2
Zewnętrzna budowa raków. A, widok grzbietowy. B, widok brzuszny.
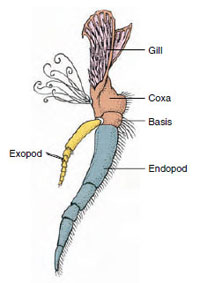
Rysunek 19-3
Części skorupiaka
dwuramiennego (trzeci maxilliped u
raków).
Położenie gonoporów zmienia się w zależności od płci i grupy skorupiaków. Mogą znajdować się na lub u podstawy pary przydatków, na końcowym końcu ciała lub na somitach bez odnóży. W raków otwory vasa deferentia są po stronie przyśrodkowej u podstawy piątej pary odnóży, a te z jajowodów są u podstawy trzeciej pary. U samic otwór do pojemnika nasiennego znajduje się zwykle w linii pośrodkowej między czwartą a piątą parą odnóży.
Apendia: Członkowie klas Malacostraca (w tym raki) i Remipedia mają zwykle parę połączonych wyrostków na każdym somicie (ryc. 19-3), chociaż somyty brzuszne w innych klasach nie mają wyrostków. Znaczna specjalizacja widoczna jest w wyrostkach pochodnych skorupiaków, takich jak raki. Wszystkie są jednak odmianami podstawowego, dwubocznego planu, zilustrowanego przez wyrostki rakowe, takie jak maxilliped (kończyna piersiowa zmodyfikowana tak, by stała się wyrostkiem głowowym) (ryc. 19-3 i 19-4). Część podstawowa, czyli protopod, posiada egzopod boczny i endopod przyśrodkowy. Protopod składa się z jednego lub dwóch stawów (basis i coxa), podczas gdy egzopod i endopod mają od jednego do kilku stawów każdy. Niektóre wyrostki, takie jak nogi chodzące u raków, stały się wtórnie uniramiczne. Wyrostki przyśrodkowe lub boczne czasami występują na kończynach skorupiaków, zwane odpowiednio enditami i exitami, a exit na protopodzie nazywany jest epipodem. Epipody są często modyfikowane jako skrzela. Tabela 19-1 pokazuje, w jaki sposób różne wyrostki uległy modyfikacji w stosunku do planu biramowego, aby dopasować się do konkretnych funkcji.
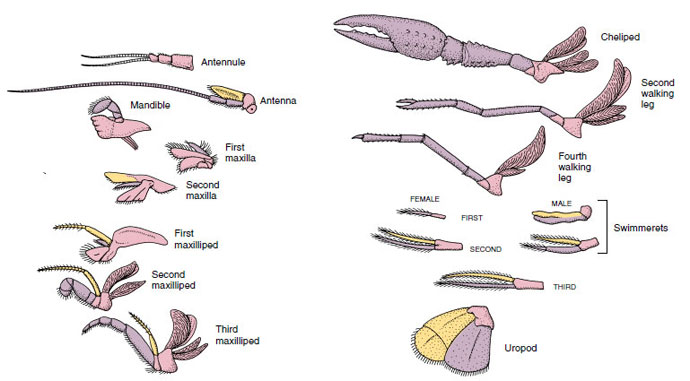
Rysunek 19-4
Apendia raka pokazujące, jak zostały zmodyfikowane z podstawowego planu biramous, jak u
swimmereta. Protopod, różowy; endopod, fioletowy; egzopod, żółty.
Struktury, które mają podobny plan podstawowy i wywodzą się ze wspólnej formy, uważa się za homologiczne, niezależnie od tego, czy pełnią tę samą funkcję, czy nie. Ponieważ wyspecjalizowane nogi chodzące, aparaty gębowe, chelipedy i swimmerety rozwinęły się ze wspólnego typu biramous, ale zostały zmodyfikowane do pełnienia różnych funkcji, wszystkie są do siebie homologiczne, co jest warunkiem znanym jako homologia seryjna. Pierwotnie kończyny były bardzo podobne, ale podczas ewolucji modyfikacji strukturalnych, niektóre gałęzie zostały zredukowane, niektóre utracone, niektóre znacznie zmienione, a niektóre nowe części dodane. Raki i ich sojusznicy posiadają najbardziej rozbudowaną homologię szeregową w królestwie zwierząt, mając 17 odrębnych, ale szeregowo homologicznych typów przydatków (Tabela 19-1).
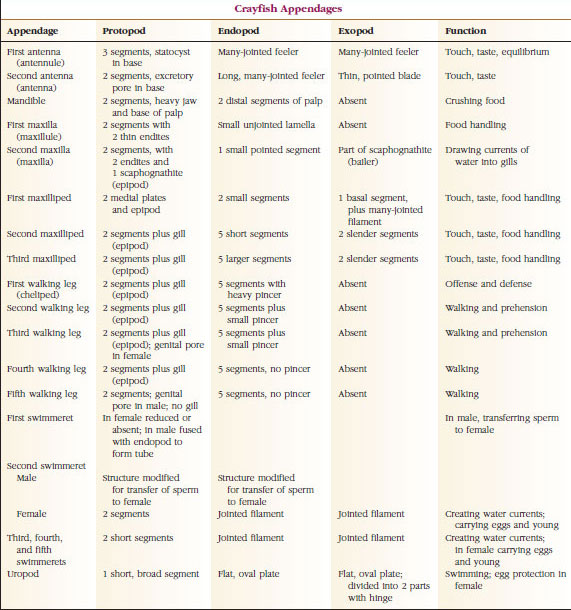
TABELA 19.1
Cechy wewnętrzne
Układ mięśniowy i nerwowy oraz segmentacja w tułowiu i odwłoku wyraźnie wykazują metameryzm odziedziczony po przodkach annelidopodobnych, ale w innych układach występują wyraźne modyfikacje. Większość zmian obejmuje koncentrację części w określonym regionie lub redukcję lub całkowitą utratę części, takich jak intersepta.
Hemocoel: Główną przestrzenią ciała u stawonogów nie jest ślimak, lecz wypełniony krwią hemocoel. Podczas rozwoju embrionalnego większości stawonogów, śladowe coelomic jamy otworzyć w mezodermie co najmniej niektórych somitów. Wkrótce ulegają one zatarciu lub stają się ciągłe z przestrzenią między rozwijającymi się strukturami mezodermalnymi i ektodermalnymi a żółtkiem. Przestrzeń ta staje się hemocoelem, a więc nie jest wyścielona przez mezodermalną otrzewną. U skorupiaków jedynymi pozostałymi przedziałami koelomicznymi są końcowe woreczki narządów wydalniczych i przestrzeń wokół gonad.
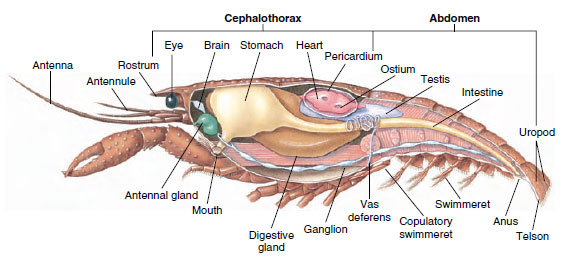
Układ mięśniowy: Mięśnie prążkowane stanowią znaczną część ciała większości Crustacea. Mięśnie są zwykle ułożone w grupach antagonistycznych: zginacze, które przyciągają część w kierunku ciała i rozciągacze, które rozciągają go na zewnątrz. Odwłok raka ma silne zginacze (Rysunek 19-5), które są wykorzystywane, gdy zwierzę płynie do tyłu, co jest jego najlepszą drogą ucieczki. Silne mięśnie po obu stronach żołądka kontrolują żuchwy.
Rysunek 19-5
Wewnętrzna budowa samca raka.
Układ oddechowy: Wymiana gazowa układu oddechowego u mniejszych skorupiaków zachodzi na cieńszych obszarach kutikuli (na przykład na odnóżach) lub w całym ciele, a wyspecjalizowane struktury mogą być nieobecne. Większe skorupiaki mają skrzela, które są delikatnymi, pierzastymi występami o bardzo cienkim kutikuliku. U dziesięcioraczków boki pancerza zamykają jamę skrzelową, która jest otwarta przednio i brzusznie (rysunek 19-6). Skrzela mogą wystawać ze ściany opłucnej do jamy skrzelowej, z połączenia odnóży piersiowych z ciałem lub z gruczołów piersiowych. Te dwa ostatnie typy są typowe dla raków. Bailer”, część drugiej szczęki, przeciąga wodę przez włókna skrzelowe, do jamy skrzelowej u podstawy odnóży i z jamy skrzelowej w przedniej części.
Układ krążenia: Skorupiaki i inne stawonogi mają „otwarty” lub lakunarny typ układu krążenia. Oznacza to, że nie ma żył i nie ma oddzielenia krwi od płynu śródmiąższowego, jak to ma miejsce u zwierząt z układem zamkniętym. Hemolimfa (krew) opuszcza serce za pomocą tętnic, krąży przez hemocel i wraca do zatok żylnych lub przestrzeni, zamiast do żył, zanim ponownie znajdzie się w sercu. Annelidae mają system zamknięty, podobnie jak kręgowce.
Grzbietowe serce jest głównym organem napędowym. Jest to jednokomorowy worek z mięśni prążkowanych. Hemolimfa dostaje się do serca z otaczającej je zatoki osierdziowej przez sparowane ujścia, z zastawkami zapobiegającymi cofaniu się do zatoki (Rysunek 19-6). Z serca hemolimfa dostaje się do jednej lub więcej tętnic. Zastawki w tętnicach zapobiegają cofaniu się hemolimfy. Małe tętnice opróżniają się do zatok tkankowych, które z kolei często uchodzą do dużej zatoki mostkowej (rysunek 19-6).
Stamtąd dośrodkowe kanały zatokowe przenoszą hemolimfę do skrzeli, jeśli są obecne, w celu wymiany tlenu i dwutlenku węgla. Hemolimfa wraca następnie kanałami eferentnymi do zatoki osierdziowej (rysunek 19-6).
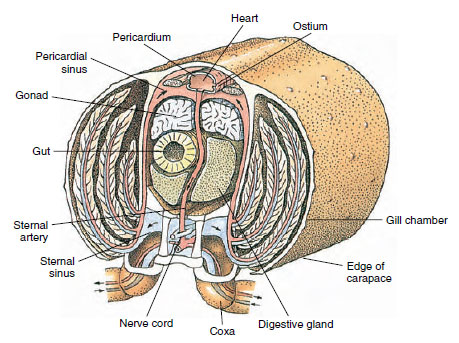
Rysunek 19-6
Diagramatyczny przekrój przez okolicę serca raka przedstawiający kierunek przepływu krwi w tym „otwartym”
układzie krwionośnym. Serce pompuje krew do tkanek ciała przez tętnice, które opróżniają się do zatok tkankowych. Powracająca
krew wchodzi do zatoki mostkowej, następnie przechodzi przez skrzela w celu wymiany gazowej, a w końcu wraca do zatoki osierdziowej przez kanały eferentne
. Należy zwrócić uwagę na brak żył.
Hemolimfa u stawonogów jest w dużej mierze bezbarwna. Obejmuje ona komórki ameboidalne co najmniej dwóch typów. Hemocyjanina, pigment oddechowy zawierający miedź, lub hemoglobina, pigment zawierający żelazo, mogą być przenoszone w roztworze. Hemolimfa ma właściwość krzepnięcia, co zapobiega jej utracie przy drobnych urazach. Niektóre komórki ameboidalne wydzielają koagulant podobny do trombiny, który wytrąca skrzep.
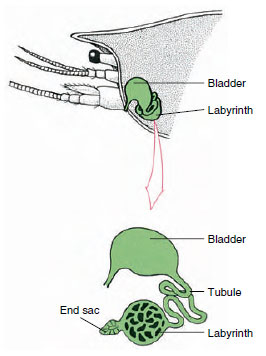
Ryc. 19-7
Schemat gruczołu antenowego (zielonego gruczołu) u
raków. (Niektóre skorupiaki nie mają labiryntu, a przewód wydalniczy (kanał nefrydialny) jest
dużo zwiniętą rurką.
Układ wydalniczy: Narządy wydalnicze dorosłych skorupiaków to para rurkowatych struktur znajdujących się w brzusznej części głowy przed przełykiem (Rysunek 19-5). Nazywane są gruczołami antenalnymi lub szczękowymi, w zależności od tego, czy otwierają się u podstawy czułków, czy u drugiej szczęki. U nielicznych dorosłych skorupiaków występują oba rodzaje. Narządami wydalniczymi dziesięcionogów są gruczoły antenowe, zwane też w tej grupie gruczołami zielonymi. Skorupiaki nie mają kanalików Malpighiego, narządów wydalniczych pająków i owadów.
Worek końcowy gruczołu antenowego, który pochodzi z embrionalnego przedziału koelomicznego, składa się z małej pęcherzyka (saccule) i gąbczastej masy zwanej labiryntem. Labirynt łączy się kanalikiem wydalniczym z grzbietowym pęcherzem moczowym, który otwiera się na zewnątrz przez por na brzusznej powierzchni podstawowego segmentu antenowego (rysunek 19-7). Ciśnienie hydrostatyczne w hemocelu zapewnia siłę do filtracji płynu do worka końcowego. Gdy filtrat przechodzi przez kanaliki wydalnicze i pęcherz moczowy, jest modyfikowany przez resorpcję soli, aminokwasów, glukozy i wody, a w końcu wydalany jako mocz.
Wydalanie odpadów azotowych (głównie amoniaku) odbywa się przez dyfuzję przez cienkie obszary kutikuli, zwłaszcza skrzeli, a tak zwane narządy wydalnicze funkcjonują głównie w celu regulacji składu jonowego i osmotycznego płynów ustrojowych. Skorupiaki słodkowodne, takie jak raki, są stale zagrożone nadmiernym rozcieńczeniem ich krwi przez wodę, która dyfunduje przez skrzela i inne powierzchnie przepuszczalne dla wody. Zielone gruczoły, tworząc rozcieńczony, niskosolny mocz, działają jako skuteczne urządzenie „kontrolujące powódź”. Niektóre Na+ i Cl- są tracone w moczu, ale ta strata jest kompensowana przez aktywną absorpcję rozpuszczonej soli przez skrzela. U skorupiaków morskich, takich jak homary i kraby, nerki pełnią funkcję regulującą skład soli w hemolimfie poprzez selektywną modyfikację zawartości soli w moczu kanalikowym. W tych formach mocz pozostaje izosmotyczny w stosunku do krwi.
Układy nerwowe i sensoryczne: Układy nerwowe skorupiaków i parzydełkowców mają wiele wspólnego, chociaż te u skorupiaków mają więcej zwojów nerwowych (rysunek 19-5). Mózg to para zwojów nadprzełykowych, które zaopatrują w nerwy oczy i dwie pary czułków. Jest on połączony łącznikami ze zwojem podprzełykowym, fuzją co najmniej pięciu par zwojów, które zaopatrują w nerwy jamę gębową, przydatki, przełyk i gruczoły antenowe. Podwójny brzuszny przewód nerwowy ma parę zwojów dla każdego somitu i nerwów obsługujących przydatki, mięśnie i inne części.
Oprócz tego centralnego układu, może istnieć współczulny układ nerwowy związany z przewodem pokarmowym.
Skorupiaki mają lepiej rozwinięte narządy zmysłów niż małże. Największymi narządami zmysłów u raków są oczy i statocysty. Szeroko rozmieszczone na ciele są dotykowe włosy, delikatne projekcje kutikuli, które są szczególnie obfite na chelae, narządów gębowych i telson. Chemiczne zmysły smaku i zapachu znajdują się we włoskach na czułkach, narządach gębowych i innych miejscach.
Podobna do worka statocysta, otwierająca się na powierzchnię przez grzbietowy por, znajduje się na podstawowym segmencie każdej pierwszej anteny raków. Statocysta zawiera grzbiet, na którym znajdują się włoski czuciowe utworzone z chitynowej wyściółki i ziaren piasku, które służą jako statolity. Kiedy zwierzę zmienia swoją pozycję, odpowiednie zmiany w położeniu ziaren na włoskach czuciowych są przekazywane jako bodźce do mózgu, a zwierzę może się odpowiednio dostosować. Każda zmiana kutikuli (ekdeza) powoduje utratę kutikularnej wyściółki statocysty, a wraz z nią ziaren piasku. Nowe ziarna są pobierane przez por grzbietowy po ekdyzie.
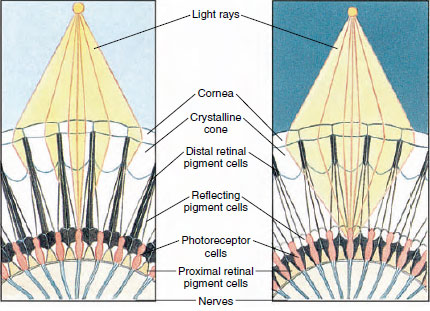
Rysunek 19-8
Porcja oka złożonego stawonoga ukazująca migrację pigmentu
w ommatidiach dla widzenia w dzień i w nocy. Pięć ommatidiów jest przedstawionych na
każdym diagramie. W dzień każde ommatidium jest otoczone ciemnym
kolorem pigmentu, tak że każde ommatidium jest stymulowane tylko przez promienie świetlne
wchodzące do jego własnej rogówki (widzenie mozaikowe); w nocy pigment tworzy
niekompletne kola i promienie świetlne mogą rozprzestrzeniać się na sąsiednie ommatidia
(obraz ciągły lub superpozycja).
Oczy u wielu skorupiaków są złożone, składają się z wielu jednostek fotoreceptorów zwanych ommatidiami (Rysunek 19-8). Zaokrągloną powierzchnię każdego oka pokrywa przezroczysty obszar kutikuli, rogówka, która jest podzielona na wiele małych kwadratów lub sześciokątów, zwanych fasetami. Fasety te są zewnętrznymi końcami ommatidiów. Każde ommatidium zachowuje się jak maleńkie oko i zawiera kilka rodzajów komórek ułożonych w sposób kolumnowy (Rysunek 19-8). Czarne komórki pigmentowe znajdują się pomiędzy sąsiednimi ommatidiami.
Ruch pigmentu w oku złożonym stawonogów pozwala mu na dostosowanie się do różnych ilości światła. Istnieją trzy zestawy komórek pigmentowych w każdym ommatidium: dystalny siatkówki, proksymalny siatkówki, i odbijające; są one tak ułożone, że mogą tworzyć mniej lub bardziej kompletny kołnierz lub rękaw wokół każdego ommatidium. W przypadku silnego oświetlenia lub adaptacji dziennej pigment siatkówki dystalnej przesuwa się do wewnątrz i spotyka się z przesuwającym się na zewnątrz pigmentem siatkówki proksymalnej, tak że wokół ommatidium tworzy się kompletny rękaw pigmentowy (Rysunek 19-8). W tym stanie tylko promienie, które uderzają bezpośrednio w rogówkę, dotrą do komórek fotoreceptorowych (siatkówkowych), ponieważ każde ommatidium jest osłonięte od innych. W ten sposób każde ommatidium widzi tylko ograniczony obszar pola widzenia (obraz mozaikowy lub apozycyjny). W słabym świetle pigmenty dystalne i proksymalne rozdzielają się, tak że promienie świetlne, przy pomocy odbijających je komórek pigmentowych, mają szansę rozprzestrzenić się na sąsiednie ommatidia i utworzyć ciągły, lub superpozycyjny, obraz. Ten drugi typ widzenia jest mniej precyzyjny, ale maksymalnie wykorzystuje ograniczoną ilość otrzymywanego światła.
Reprodukcja, cykle życiowe i funkcje endokrynologiczne
Większość skorupiaków ma oddzielne płcie i istnieją różne specjalizacje do kopulacji wśród różnych grup. Barnacles są jednopienne, ale generalnie praktykują zapłodnienie krzyżowe. W niektórych ostronosów samce są rzadkie, a reprodukcja jest zwykle partenogenetyczne. Większość skorupiaków w jakiś sposób wychowuje jaja: ramienionogi i pąkle mają specjalne komory lęgowe, widłonogi mają woreczki lęgowe przymocowane do boków odwłoka (patrz Rysunek 19-19), a wiele malakostrachów nosi jaja i młode przymocowane do wyrostków brzusznych.
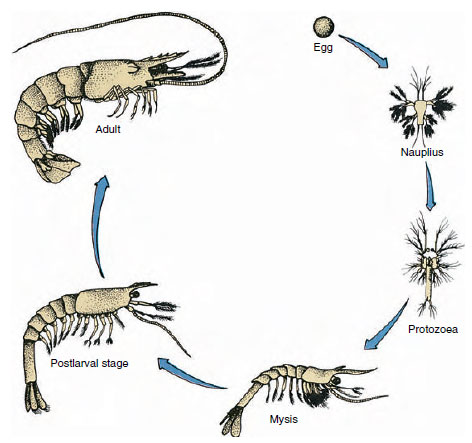
Rysunek 19-9
Cykl życiowy krewetki zatokowej Penaeus. Penaeids składają ikrę na głębokości od 40 do 90 m. Młode formy larwalne są
planktoniczne i przemieszczają się w kierunku przybrzeżnym do wód o niższym zasoleniu, gdzie rozwijają się jako osobniki młodociane. Starsze krewetki wracają do głębszych wód
na brzeg.
Raki mają rozwój bezpośredni: nie ma formy larwalnej. Z jaja wykluwa się malutki osobnik młodociany o takiej samej formie jak dorosły, z kompletnym zestawem przydatków i somitów. U większości skorupiaków rozwój jest jednak pośredni, a z jaja wykluwa się larwa zupełnie niepodobna do postaci dorosłej pod względem budowy i wyglądu. Przemiana z larwy ostatecznie w postać dorosłą to metamorfoza. Prymitywną i najczęściej występującą u skorupiaków larwą jest nauplius (rys. 19-9 i 19-23). Naupliusy mają tylko trzy pary wyrostków: jednopalczaste pierwsze antenki, dwupalczaste czułki i dwupalczaste żuchwy. Wszystkie funkcjonują na tym etapie jako wyrostki pływne.
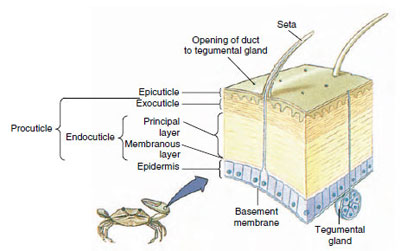
Rysunek 19-10
Struktura kutykuli skorupiaków.
Późniejszy rozwój może obejmować stopniową zmianę do postaci dorosłego ciała, a przydatki i somity są dodawane przez serię molt, lub przyjęcie postaci dorosłej może wiązać się z bardziej gwałtownymi zmianami. Na przykład metamorfoza pąkli przebiega od swobodnie pływającej nauplius do larwy z pancerzem małżowinowym zwanym cyprid i wreszcie do oseska dorosłego z wapiennymi płytkami.
Ekdeza: Ecdysis (ek´duh-sis) (Gr. ekdyein, do strip off), lub molting, jest konieczne dla ciała, aby zwiększyć rozmiar, ponieważ egzoszkielet jest nie żywy i nie rośnie jak zwierzę rośnie. Na znaczną część funkcjonowania skorupiaków, w tym na ich rozmnażanie, zachowanie i wiele procesów metabolicznych, bezpośredni wpływ ma fizjologia cyklu linienia.
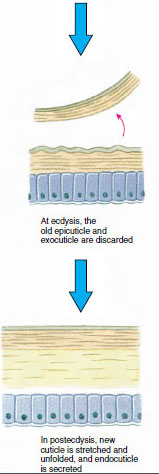
Kutikula, wydzielana przez leżący pod nią naskórek, ma kilka warstw (ryc. 19-10). Najbardziej zewnętrzną jest epikutykula, bardzo cienka warstwa białka impregnowanego lipidami. Większą część kutikuli stanowi kilka warstw proutikuli: (1) egzokutykula, która znajduje się tuż pod epikutykulą i zawiera białko, sole wapnia i chitynę; (2) endokutykula, która sama składa się z (3) warstwy zasadniczej, która zawiera więcej chityny i mniej białka i jest silnie uwapniona, oraz (4) nieskalanej warstwy błoniastej, stosunkowo cienkiej warstwy chityny i białka.
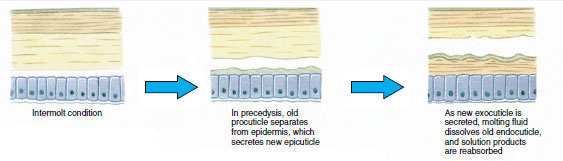
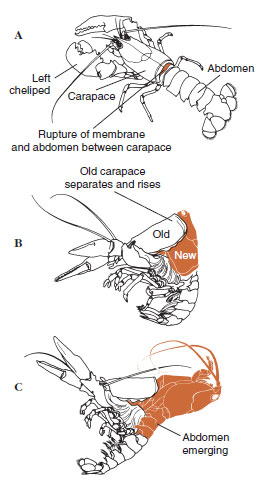
Na jakiś czas przed właściwą ekdezą komórki epidermy znacznie się powiększają. Oddzielają się one od warstwy błoniastej, wydzielają nową epikutykulę i zaczynają wydzielać nową egzokutykulę (Rysunek 19-11). Enzymy są uwalniane do obszaru powyżej nowego naskórka. Enzymy te zaczynają rozpuszczać stary endocuticle, a rozpuszczalne produkty są resorbowane i magazynowane w ciele skorupiaka. Niektóre sole wapnia są magazynowane w ścianach żołądka w postaci gastrolitów (nagromadzeń minerałów). W końcu pozostaje tylko egzokutykula i epikutykula starej kutikuli, pod którą znajduje się nowa epikutykula i nowa egzokutykula. Zwierzę połyka wodę, którą wchłania przez jelita, a objętość jego krwi znacznie wzrasta. Ciśnienie wewnętrzne powoduje rozszczepienie kutikuli i zwierzę wyrywa się ze starego egzoszkieletu (rysunek 19-12). Następnie następuje rozciąganie wciąż miękkiego nowego kutikuli, odkładanie nowej endokutikuli, ponowne odkładanie ocalałych soli nieorganicznych i innych składników oraz twardnienie nowej kutikuli. W okresie linienia zwierzę jest bezbronne i pozostaje w ukryciu.
 |
|
 |
 |
| Rysunek 19-12 Sekwencja linienia u homara, Homarus americanus. A, Błona między pancerzem a odwłokiem pęka, a pancerz zaczyna się powoli unosić. Ten etap może trwać do 2 godzin. B i C, głowa, tułów i wreszcie odwłok wycofują się. Ten proces zwykle trwa nie więcej niż 15 minut. Natychmiast po ecdysis, chelipedy są wysuszone i ciało jest bardzo miękkie. Homar kontynuuje szybkie wchłanianie wody tak, że w ciągu 12 godzin ciało zwiększa się o około 20% długości i 50% wagi. Woda w tkankach zostanie zastąpiona przez białko w kolejnych tygodniach. |
Rysunek 19-11 Wydzielanie i resorpcja skorupiaków w trakcie ekstazy. |
Gdy skorupiak jest młody, ekdiza musi występować często, aby umożliwić mu wzrost, a cykl linienia jest stosunkowo krótki. Jak zwierzę zbliża się do dojrzałości, okresy intermolt stają się stopniowo coraz dłuższe, a w niektórych gatunków molting przestaje całkowicie. Podczas okresów intermolt, wzrost masy tkanki występuje jako żywej tkanki zastępuje wody.
Hormonalna kontrola cyklu Ecdysis
Chociaż ecdysis jest hormonalnie kontrolowane, cykl jest często inicjowane przez bodziec środowiskowy postrzegane przez centralnego układu nerwowego. Takie bodźce mogą obejmować temperaturę, długość dnia i wilgotność (w przypadku krabów lądowych). Sygnał z centralnego układu nerwowego powoduje zmniejszenie produkcji hormonu hamującego linienie przez narząd X. Narząd X to grupa komórek neurosekrecyjnych w rdzeniach końcowych mózgu. U raków i innych parzystokopytnych, medulla terminalis znajduje się w gałce ocznej. Hormon jest przenoszony w aksonach organu X do gruczołu zatokowego (który sam w sobie prawdopodobnie nie jest gruczołowy w funkcji), również w gałce ocznej, gdzie jest uwalniany do hemolimfy.
spadek poziomu hormonu hamującego topnienie promuje uwalnianie hormonu topnienia z Y-organs. Yorgans są pod naskórkiem w pobliżu mięśni przywodzicieli żuchwy, i są homol-ogus do gruczołów prothoracic owadów, które produkują hormon ecdysone. Działanie hormonu rozrodczego polega na zapoczątkowaniu procesów prowadzących do ekdystezy (proekdystezy). Po zainicjowaniu, cykl przebiega automatycznie bez dalszego działania hormonów z X- lub Y-organs.
Inne funkcje endokrynne: Nie tylko usuwanie oczek przyspiesza molting, to również stwierdzono ponad 100 lat temu, że skorupiaki, których eyestalks zostały usunięte nie może już dostosować ubarwienie ciała do warunków tła. Ponad 50 lat temu odkryto, że wada ta nie jest spowodowana utratą wzroku, ale utratą hormonów w oczodołach. Kolor ciała skorupiaków jest w dużej mierze wynikiem pigmentów w specjalnych rozgałęzionych komórkach (chromatoforach) w naskórku.
Koncentracja granulek pigmentu w centrum komórek powoduje efekt rozjaśnienia, a rozproszenie pigmentu w obrębie komórek powoduje efekt przyciemnienia. Zachowanie pigmentu jest kontrolowane przez hormony z komórek neurosekrecyjnych w gałce ocznej, podobnie jak migracja pigmentu siatkówki w celu adaptacji do światła i ciemności w oczach (ryc. 19-8).
Uwolnienie materiału neurosekrecyjnego z narządów okołosercowych w ścianie osierdzia powoduje wzrost częstości i amplitudy bicia serca.
Gruczoły androgenne, po raz pierwszy znalezione u amfipoda (Orchestia, pospolity lejek plażowy), występują u samców malakostracjanów. W przeciwieństwie do większości innych narządów endokrynnych u skorupiaków, nie są to narządy neurosekrecyjne. Ich wydzielanie stymuluje ekspresję męskich cech płciowych. Młode malakostracany mają szczątkowe gruczoły androgenne, ale u samic gruczoły te nie rozwijają się. Jeśli zostaną one sztucznie wszczepione samicy, jej jajniki przekształcają się w jądra i zaczynają produkować plemniki, a przydatki zaczynają nabierać cech męskich przy następnym linieniu. U równonogów gruczoły androgenne znajdują się w jądrach; u wszystkich innych malakostracjanów znajdują się one między mięśniami ścięgien ostatnich odnóży tułowiowych i częściowo przyczepione są w pobliżu zakończeń nasieniowodów. Chociaż samice nie posiadają narządów podobnych do gruczołów androgennych, ich jajniki produkują jeden lub dwa hormony, które wpływają na drugorzędowe cechy płciowe.
Hormony, które wpływają na inne procesy zachodzące w organizmie skorupiaków mogą być obecne, a dowody sugerują, że substancja neurosekrecyjna produkowana w szyjce ocznej reguluje poziom cukru we krwi.
Zwyczaje żywieniowe
Zwyczaje żywieniowe i przystosowania do żywienia różnią się znacznie wśród skorupiaków. Wiele form może przejść z jednego typu karmienia do drugiego w zależności od środowiska i dostępności pokarmu, ale wszystkie używają tego samego podstawowego zestawu aparatów gębowych. Żuchwy i szczęki służą do połykania pokarmu; szczękoczułki chwytają i miażdżą pokarm. U drapieżników chodzące odnóża, zwłaszcza chelipedy, służą do chwytania pokarmu.
Wiele skorupiaków, zarówno dużych, jak i małych, jest drapieżnych, a niektóre mają ciekawe przystosowania do zabijania ofiar. Jedna forma krewetkopodobna, Lygiosquilla, ma na jednej z nóg chodzących wyspecjalizowaną cyfrę, która może być wciągnięta w rowek i zwolniona nagle, aby przebić przechodzącą ofiarę. Krewetki pistoletowe (Alpheus spp.) mają ogromnie powiększoną chela, która może być cocked jak młotek pistoletu i pstryknął z siłą, która ogłusza ich prey.
Pokarm karmicieli zawiesiny waha się od planktonu i detrytusu do bakterii. Drapieżniki konsumują larwy, robaki, skorupiaki, ślimaki i ryby. Scavengers jeść martwych zwierząt i roślin materii. Karmiciele zawiesinowi, tacy jak krewetki bajkalskie, pchły wodne i pąkle, wykorzystują swoje odnóża, które posiadają grube frędzle z włosków, do tworzenia prądów wodnych, które omiatają cząstki pokarmu przez włoski. Krewetki błotne (Upogebia spp.) używają długich wąsów na dwóch pierwszych parach wyrostków kolczystych do odcedzania pokarmu z wody krążącej w ich norze dzięki ruchom ich pływaków.
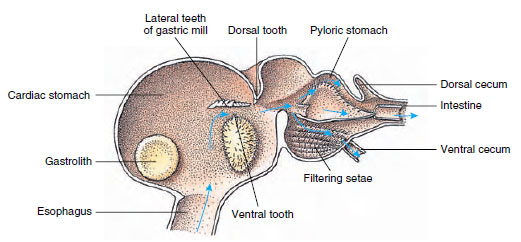
Raki mają dwuczęściowy żołądek (rysunek 19-13). W pierwszej części znajduje się młynek żołądkowy, w którym pokarm, już rozdrobniony przez żuchwy, może być dalej rozdrobniony przez trzy zęby wapienne na cząstki wystarczająco drobne, aby przejść przez filtr z setozy w drugiej części; cząstki pokarmu przechodzą następnie do jelita w celu trawienia chemicznego.
Ryc. 19-13
Żołądek malakostracjana ukazujący „młynek” żołądkowy i kierunki ruchów pokarmu. Młynek ma chitynowe grzbiety lub
zęby, służące do żucia, oraz włoski do odcedzania pokarmu przed przejściem do odźwiernika żołądka.
Dodaj komentarz