Uwalnianie neuroprzekaźników
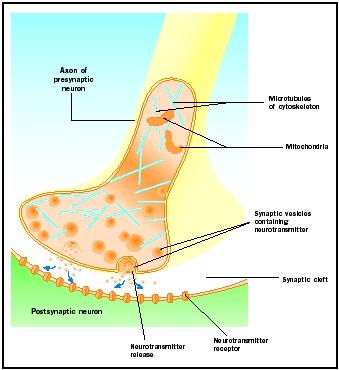
Potencjały czynnościowe docierające do gałek synaptycznych wyzwalają uwalnianie neuroprzekaźnika do szczeliny synaptycznej. Mechanizm molekularny nie jest do końca poznany. Między nadejściem potencjału czynnościowego a uwolnieniem neuroprzekaźnika występuje „opóźnienie synaptyczne” wynoszące od jednej do dwóch milisekund. Potencjały czynnościowe otwierają kanały wapniowe w błonie gałki synaptycznej, co powoduje ruch jonów wapnia do wewnątrz. Jony wapnia wyzwalają uwalnianie neuroprzekaźnika z pęcherzyków synaptycznych do szczeliny synaptycznej. Pęcherzyki synaptyczne łączą się z błoną presynaptyczną podczas tego procesu egzocytozy. Błony starych pęcherzyków stają się częścią błony presynaptycznej, a nowe pęcherzyki odrywają się od przylegającego obszaru błony. Te nowe pęcherzyki są następnie ponownie wypełnione nowo zsyntetyzowanymi lub „przetworzonymi” neuroprzekaźnikami.
Uwolnione neuroprzekaźniki dyfundują przez wąską szczelinę synaptyczną. Na błonie postsynaptycznej, cząsteczki neuroprzekaźnika wiążą się z cząsteczkami receptora związanymi z błoną z miejscami rozpoznawania specyficznymi dla tego neuroprzekaźnika. Wiązanie neuroprzekaźnika z receptorem wyzwala odpowiedź postsynaptyczną specyficzną dla tego receptora. Odpowiedzi te mogą mieć charakter pobudzający lub hamujący, w zależności od właściwości receptora. Jeśli stymulacja receptora powoduje, że błona postsynaptyczna staje się bardziej elektrycznie dodatnia (zdepolaryzowana), jest to pobudzający potencjał postsynaptyczny (EPSP). Jeśli jest on bardziej ujemny (hiperpolaryzowany), jest to hamujący potencjał postsynaptyczny (IPSP). Pobudzenie i hamowanie zależy od właściwości receptora, a nie od neuroprzekaźnika. Receptory sprzężone z kanałami sodowymi lub wapniowymi są pobudzające i powodują depolaryzację błony postsynaptycznej, natomiast receptory sprzężone z kanałami chlorkowymi lub potasowymi są hamujące i powodują hiperpolaryzację błony postsynaptycznej. Takie receptory sprzężone z kanałami jonowymi nazywane są receptorami jonotropowymi.
Inne receptory są sprzężone z systemami „drugiego przekaźnika”, które inicjują serię reakcji biochemicznych w komórce postsynaptycznej. Są to receptory metabotropowe . Receptory metabotropowe mogą wytwarzać wiele różnych
zdarzenia postsynaptyczne. Obejmują one zakres od bezpośredniej aktywacji sąsiednich kanałów jonowych, do zmiany wrażliwości receptora, do transkrypcji określonych kwasów rybonukleinowych (RNA), a nawet aktywacji określonych genów . Synapsy chemiczne są częścią bardzo adaptowalnego i elastycznego systemu komunikacji. Nie są to statyczne struktury anatomiczne o stałych właściwościach, ale są to struktury dynamiczne, zdolne do zmiany swoich właściwości molekularnych w zmieniających się okolicznościach.
Istnieją dosłownie setki neurotransmiterów. Niektóre z nich to dość proste związki, takie jak acetylocholina, serotonina, katecholaminy (dopamina, noradrenalina i epinefryna) oraz szereg aminokwasów. Wiele z nich jest bardziej złożonych i należy do szerokiej gamy przekaźników neuropeptydowych. Po uwolnieniu do szczeliny synaptycznej neuroprzekaźniki pozostają aktywne, dopóki nie zostaną zmienione chemicznie lub zabrane z powrotem do gałki synaptycznej przez specjalne systemy nośników i poddane recyklingowi. W synapsach cholinergicznych, w szczelinie synaptycznej obecna jest acetylocholinoesteraza. Enzym ten rozszczepia neuroprzekaźnik na octan i cholinę, z których żaden nie jest aktywny. Z kolei serotonina i epinefryna są pobierane do terminala presynaptycznego i poddawane recyklingowi.
Dodaj komentarz