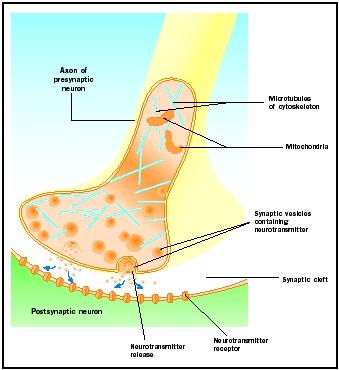
Rilascio del neurotrasmettitore
I potenziali d’azione che arrivano ai pomelli sinaptici innescano il rilascio del neurotrasmettitore nella fessura sinaptica. Il meccanismo molecolare non è completamente compreso. Un “ritardo sinaptico” di uno o due millisecondi si verifica tra l’arrivo del potenziale d’azione e il rilascio del neurotrasmettitore. I potenziali d’azione aprono i canali del calcio nella membrana della manopola sinaptica, che provoca un movimento verso l’interno di ioni calcio . Gli ioni di calcio innescano il rilascio del neurotrasmettitore dalle vescicole sinaptiche nella fessura sinaptica. Le vescicole sinaptiche si fondono con la membrana presinaptica durante questo processo di esocitosi. Le membrane delle vecchie vescicole diventano parte della membrana presinaptica e nuove vescicole si staccano da una zona adiacente della membrana. Queste nuove vescicole sono successivamente riempite di neurotrasmettitori appena sintetizzati o “riciclati”.
I neurotrasmettitori rilasciati si diffondono attraverso la stretta fessura sinaptica. Sulla membrana postsinaptica, le molecole di neurotrasmettitore si legano a molecole di recettori legati alla membrana con siti di riconoscimento specifici per quel neurotrasmettitore. Il legame del neurotrasmettitore al recettore innesca una risposta postsinaptica specifica per quel recettore. Queste risposte possono essere eccitatorie o inibitorie, a seconda delle proprietà del recettore. Se la stimolazione del recettore fa sì che la membrana postsinaptica diventi elettricamente più positiva (depolarizzata), si tratta di un potenziale postsinaptico eccitatorio (EPSP). Se è più negativo (iperpolarizzato), si tratta di un potenziale postsinaptico inibitorio (IPSP). L’eccitazione e l’inibizione dipendono dalle proprietà del recettore e non dal neurotrasmettitore. I recettori accoppiati ai canali del sodio o del calcio sono eccitatori e producono una depolarizzazione della membrana postsinaptica, mentre i recettori accoppiati ai canali del cloruro o del potassio sono inibitori e producono una iperpolarizzazione della membrana postsinaptica. Tali recettori accoppiati ai canali ionici sono chiamati recettori ionotropi.
Altri recettori sono accoppiati a sistemi di “secondo messaggero” che iniziano una serie di reazioni biochimiche nella cellula postsinaptica. Questi sono recettori metabotropi. I recettori metabotropi possono produrre molti diversi
eventi postsinaptici. Questi vanno dall’attivazione diretta dei canali ionici adiacenti, all’alterazione della sensibilità del recettore, alla trascrizione di specifici acidi ribonucleici messaggeri (RNA), o anche l’attivazione di geni specifici. Le sinapsi chimiche fanno parte di un sistema di comunicazione molto adattabile e flessibile. Non si tratta di strutture anatomiche statiche con proprietà fisse, ma di strutture dinamiche, in grado di cambiare le loro proprietà molecolari al variare delle circostanze.
Ci sono letteralmente centinaia di neurotrasmettitori. Alcuni sono composti abbastanza semplici come l’acetilcolina, la serotonina, le catecolamine (dopamina, norepinefrina ed epinefrina) e un certo numero di aminoacidi. Molti sono più complessi e appartengono alla vasta gamma di trasmettitori neuropeptidici. Una volta rilasciati nella fessura sinaptica, i neurotrasmettitori rimangono attivi finché non vengono alterati chimicamente o riportati nella manopola sinaptica da speciali sistemi di trasporto e riciclati. Nelle sinapsi colinergiche, l’acetilcolinesterasi è presente nella fessura sinaptica. Questo enzima scinde il neurotrasmettitore in acetato e colina, nessuno dei quali è attivo. La serotonina e l’epinefrina, invece, sono assorbite nel terminale presinaptico e riciclate.
Lascia un commento