Subphylum Crustacea
Natura generale di un crostaceo
I crostacei differiscono dagli altri artropodi in vari modi, ma l’unica caratteristica veramente distintiva è che i crostacei sono gli unici artropodi con due coppie di antenne. Oltre a due paia di antenne e un paio di mandibole, i crostacei hanno due paia di mascelle sulla testa, seguite da un paio di appendici su ogni segmento del corpo o somite. In alcuni crostacei non tutti i somiti portano appendici. Tutte le appendici, tranne forse le prime antenne, sono primitivamente biramose (due rami principali), e almeno alcune appendici degli adulti attuali mostrano questa condizione. Gli organi specializzati per la respirazione, se presenti, funzionano come branchie.
La maggior parte dei crostacei ha tra 16 e 20 somiti, ma alcune forme hanno 60 somiti o più. Un numero maggiore di somiti è una caratteristica primitiva. La condizione più derivata è quella di avere meno segmenti e una maggiore tagmatizzazione. I principali tagmata sono la testa, il torace e l’addome, ma questi non sono omologhi in tutta la classe (o anche all’interno di alcune sottoclassi) a causa dei diversi gradi di fusione dei somiti, per esempio, come nel cefalotorace.
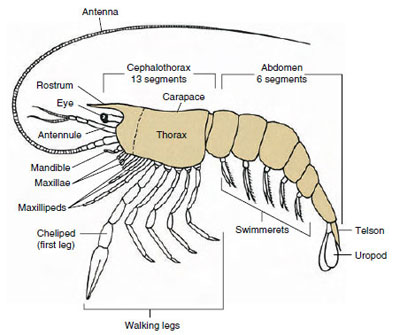
Figura 19-1
Piano archetipico dei Malacostraca. Le due mascelle e i tre
massillipedi sono stati separati diagrammaticamente per illustrare il
piano generale.
Il gruppo di gran lunga più grande di crostacei è la classe Malacostraca, che comprende aragoste, granchi, gamberi, saltatori da spiaggia, cimici, e molti altri. Questi mostrano una disposizione sorprendentemente costante dei segmenti del corpo e dei tagmata, che è considerata la pianta ancestrale della classe (Figura 19-1). Questo tipico piano corporeo ha una testa di cinque somiti (sei embrionalmente) fusi, un torace di otto somiti e un addome di sei somiti (sette in alcune specie). All’estremità anteriore c’è un rostro non segmentato e all’estremità posteriore c’è un telson non segmentato, che con l’ultimo somite addominale e i suoi uropodi forma un ventaglio di coda in molte forme.
In molti crostacei la cuticola dorsale della testa può estendersi posteriormente e intorno ai lati dell’animale per coprire o essere fusa con alcuni o tutti i somiti toracici e addominali. Questa copertura è chiamata carapace. In alcuni gruppi il carapace forma valvole simili a conchiglie che coprono la maggior parte o tutto il corpo. Nei decapodi (tra cui aragoste, gamberi, granchi e altri), il carapace copre l’intero cefalotorace ma non l’addome.
Forma e funzione
A causa delle loro dimensioni e facile reperibilità, i grandi crostacei come i gamberi sono stati studiati più di altri gruppi. Sono anche comunemente studiati nei corsi di laboratorio introduttivi. Pertanto molti dei commenti che seguono si applicano specificamente ai gamberi e ai loro parenti.
Caratteristiche esterne
I corpi dei crostacei sono coperti da una cuticola secreta composta da chitina, proteine e materiale calcareo. Le placche più dure e pesanti dei crostacei più grandi sono particolarmente ricche di depositi calcarei. Il duro rivestimento protettivo è morbido e sottile in corrispondenza delle articolazioni tra i somiti, permettendo la flessibilità dei movimenti. Il carapace, se presente, copre gran parte o tutto il cefalotorace; nei decapodi come i gamberi, tutti i segmenti della testa e del torace sono racchiusi dorsalmente dal carapace. Ogni somite non racchiuso dal carapace è coperto da una piastra cuticolare dorsale, o tergum (Figura 19-2A), e una barra trasversale ventrale, lo sterno, si trova tra le appendici segmentali (Figura 19-2B). L’addome termina in un telson, che non è considerato un somite e porta l’ano. (Il telson può essere omologo al pigidio degli anellidi)
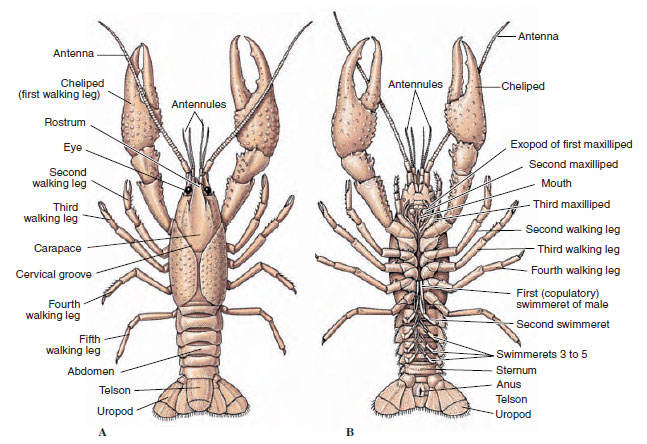
Figura 19-2
Struttura esterna dei gamberi. A, vista dorsale. B, vista ventrale.
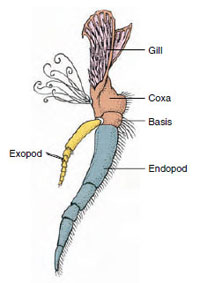
Figura 19-3
Parti di un crostaceo biramoso
appendice (terzo maxillipede di un
gambero).
La posizione dei gonopori varia secondo il sesso e il gruppo di crostacei. Possono trovarsi su o alla base di un paio di appendici, all’estremità terminale del corpo, o su somiti senza gambe. Nei gamberi le aperture dei vasa deferentia sono sul lato mediano alla base del quinto paio di zampe, e quelle degli ovidotti sono alla base del terzo paio. Nelle femmine l’apertura del ricettacolo seminale si trova di solito nella linea medioventrale tra il quarto e il quinto paio di zampe mobili.
Appendici: I membri delle classi Malacostraca (compresi i gamberi) e Remipedia hanno tipicamente un paio di appendici articolate su ogni somite (Figura 19-3), anche se i somiti addominali nelle altre classi non portano appendici. Una notevole specializzazione è evidente nelle appendici dei crostacei derivati come i gamberi. Tuttavia, tutte sono variazioni del piano di base, biramoso, illustrato da un’appendice del gambero come il maxillipede (un arto toracico modificato per diventare un’appendice della testa) (Figure 19-3 e 19-4). La porzione basale, o protopode, porta un esopode laterale e un endopode mediale. Il protopode è composto da una o due articolazioni (base e coxa), mentre l’esopode e l’endopode hanno da una a più articolazioni ciascuno. Alcune appendici, come le zampe ambulanti dei gamberi, sono diventate secondariamente univoche. Processi mediali o laterali si verificano a volte sugli arti dei crostacei, chiamati rispettivamente enditi ed esiti, e un esite sul protopode è chiamato epipode. Gli epipodi sono spesso modificati come branchie. La tabella 19-1 mostra come le varie appendici si sono modificate dal piano biramidale per adattarsi a funzioni specifiche.
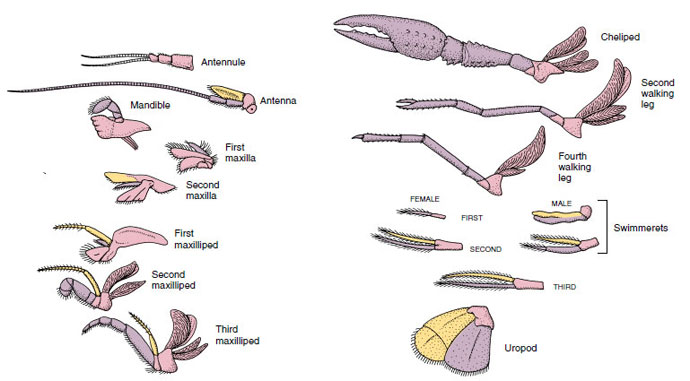
Figura 19-4
Appendici di un gambero di fiume che mostrano come sono stati modificati dalla pianta biramosa di base, come in un
gambero. Protopode, rosa; endopode, viola; esopode, giallo.
Strutture che hanno un piano di base simile e che sono discese da una forma comune sono dette omologhe, che abbiano o meno la stessa funzione. Poiché le gambe specializzate per camminare, l’apparato boccale, i chelipedi e i nuotatori si sono tutti sviluppati da un tipo biramoso comune, ma sono stati modificati per svolgere funzioni diverse, sono tutti omologhi tra loro, una condizione nota come omologia seriale. Primitivamente gli arti erano tutti molto simili, ma durante l’evoluzione delle modifiche strutturali, alcuni rami sono stati ridotti, alcuni persi, alcuni notevolmente alterati, e alcune nuove parti aggiunte. I gamberi e i loro alleati possiedono l’omologia seriale più elaborata del regno animale, avendo 17 tipi di appendici distinte ma omologate in serie (Tabella 19-1).
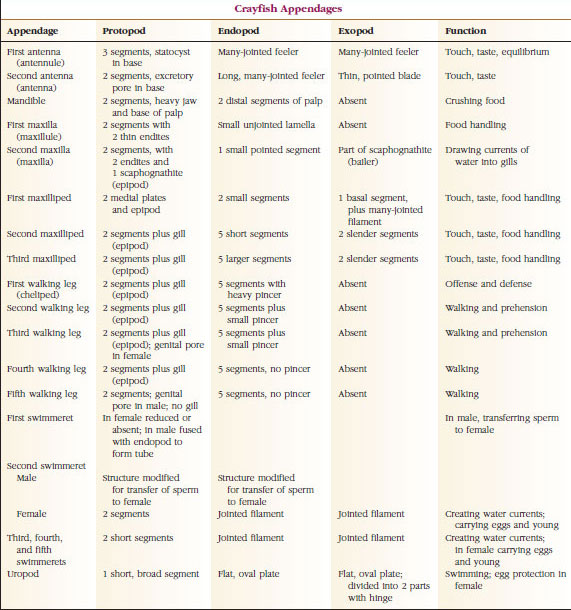
Tabella 19.1
Caratteristiche interne
I sistemi muscolare e nervoso e la segmentazione nel torace e nell’addome mostrano chiaramente un metamerismo ereditato da antenati simili agli anellidi, ma ci sono marcate modifiche in altri sistemi. La maggior parte dei cambiamenti comporta la concentrazione di parti in una regione particolare o la riduzione o la perdita completa di parti, come l’intersepta.
Hemocoel: Lo spazio corporeo principale negli artropodi non è il celoma ma un emoceleste pieno di sangue. Durante lo sviluppo embrionale della maggior parte degli artropodi, cavità celomiche vestigiali si aprono nel mesoderma di almeno alcuni somiti. Queste sono presto obliterate o diventano continue con lo spazio tra le strutture mesodermiche ed ectodermiche in via di sviluppo e il tuorlo. Questo spazio diventa l’emocele e non è quindi rivestito da un peritoneo mesodermico. Nei crostacei gli unici compartimenti celomici rimasti sono i sacchi terminali degli organi escretori e lo spazio intorno alle gonadi.
Sistema muscolare: I muscoli striati costituiscono una parte considerevole del corpo della maggior parte dei crostacei. I muscoli sono di solito disposti in gruppi antagonisti: flessori, che attirano una parte verso il corpo, ed estensori, che la estendono verso l’esterno. L’addome di un gambero ha potenti flessori (Figura 19-5), che sono usati quando l’animale nuota all’indietro – il suo miglior mezzo di fuga. Forti muscoli su entrambi i lati dello stomaco controllano le mandibole.
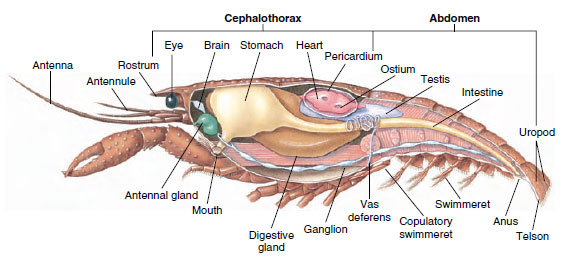
Figura 19-5
Struttura interna di un gambero maschio.
Sistema respiratorio: Lo scambio di gas respiratorio nei crostacei più piccoli avviene su aree più sottili di cuticola (per esempio, nelle gambe) o sull’intero corpo, e le strutture specializzate possono essere assenti. I crostacei più grandi hanno le branchie, che sono delicate, proiezioni simili a piume con cuticola molto sottile. Nei decapodi i lati del carapace racchiudono la cavità branchiale, che è aperta anteriormente e ventralmente (Figura 19-6). Le branchie possono sporgere dalla parete pleurica nella cavità branchiale, dall’articolazione delle gambe toraciche con il corpo, o dalle coxe toraciche. Gli ultimi due tipi sono tipici dei gamberi. Il “bailer”, una parte della seconda mascella, attira l’acqua sui filamenti branchiali, nella cavità branchiale alla base delle zampe, e fuori dalla cavità branchiale nella parte anteriore.
Sistema circolatorio: I crostacei e altri artropodi hanno un sistema circolatorio di tipo “aperto” o lacunare. Questo significa che non ci sono vene e nessuna separazione del sangue dal fluido interstiziale, come avviene negli animali con sistemi chiusi. L’emolinfa (sangue) lascia il cuore attraverso le arterie, circola attraverso l’emocoel e ritorna nei seni venosi, o spazi, invece che nelle vene, prima di rientrare nel cuore. Gli anellidi hanno un sistema chiuso, come i vertebrati.
Un cuore dorsale è il principale organo propulsivo. È un sacco monocamerale di muscoli striati. L’emolinfa entra nel cuore dal seno pericardico circostante attraverso ostia accoppiati, con valvole che impediscono il reflusso nel seno (Figura 19-6). Dal cuore l’emolinfa entra in una o più arterie. Le valvole nelle arterie impediscono il riflusso dell’emolinfa. Le piccole arterie si svuotano nei seni tissutali, che a loro volta spesso scaricano in un grande seno sternale (Figura 19-6).
Da lì, i canali afferenti del seno portano l’emolinfa alle branchie, se presenti, per lo scambio di ossigeno e anidride carbonica. L’emolinfa ritorna poi al seno pericardico attraverso i canali efferenti (Figura 19-6).
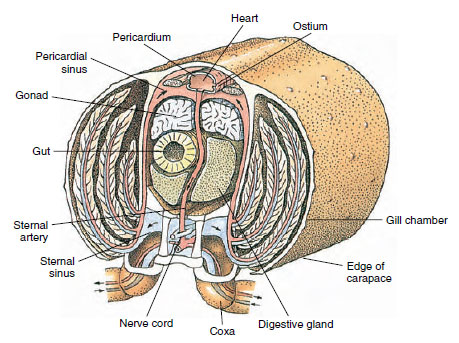
Figura 19-6
Sezione trasversale di un gambero che mostra la direzione del flusso sanguigno in questo sistema “aperto”
sangue. Il cuore pompa il sangue ai tessuti del corpo attraverso le arterie, che si svuotano nei seni dei tessuti. Il sangue
di ritorno entra nel seno sternale, poi passa attraverso le branchie per lo scambio di gas, e infine torna al seno pericardico attraverso i canali efferenti
. Si noti l’assenza di vene.
L’emolinfa negli artropodi è in gran parte incolore. Comprende cellule ameboidi di almeno due tipi. L’emocianina, un pigmento respiratorio contenente rame, o l’emoglobina, un pigmento contenente ferro, possono essere trasportati in soluzione. L’emolinfa ha la proprietà di coagulare, il che impedisce la sua perdita nelle lesioni minori. Alcune cellule ameboidi rilasciano un coagulante simile alla trombina che precipita la coagulazione.
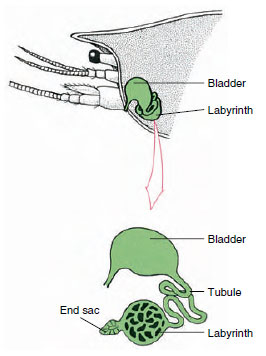
Figura 19-7
Schema della ghiandola antennale (ghiandola verde) dei
grayfish. (Alcuni crostacei mancano di un labirinto,
e il tubulo escretore (canale nefridiale) è
un tubo molto arrotolato.
Sistema escretore: Gli organi escretori dei crostacei adulti sono un paio di strutture tubolari situate nella parte ventrale della testa davanti all’esofago (Figura 19-5). Sono chiamate ghiandole antennali o ghiandole mascellari, a seconda che si aprano alla base delle antenne o delle seconde mascelle. Alcuni crostacei adulti le hanno entrambe. Gli organi escretori dei decapodi sono le ghiandole antennali, chiamate anche ghiandole verdi in questo gruppo. I crostacei non hanno tubuli malpighiani, gli organi escretori dei ragni e degli insetti.
Il sacco terminale della ghiandola antennale, che deriva da un compartimento celomico embrionale, consiste in una piccola vescicola (saccula) e una massa spugnosa chiamata labirinto. Il labirinto si collega tramite un tubulo escretore a una vescica dorsale, che si apre all’esterno tramite un poro sulla superficie ventrale del segmento antennale basale (Figura 19-7). La pressione idrostatica all’interno dell’emocoel fornisce la forza per la filtrazione del fluido nel sacco terminale. Mentre il filtrato passa attraverso il tubulo escretore e la vescica, viene modificato dal riassorbimento di sali, aminoacidi, glucosio e un po’ d’acqua e viene infine escreto come urina.
L’escrezione dei rifiuti azotati (soprattutto ammoniaca) avviene per diffusione attraverso aree sottili di cuticola, specialmente le branchie, e i cosiddetti organi escretori funzionano principalmente per regolare la composizione ionica e osmotica dei fluidi corporei. I crostacei d’acqua dolce, come i gamberi, sono costantemente minacciati dalla sovradiluizione del loro sangue da parte dell’acqua, che si diffonde attraverso le branchie e altre superfici permeabili all’acqua. Le ghiandole verdi, formando un’urina diluita e a basso contenuto di sale, agiscono come un efficace dispositivo di “controllo delle inondazioni”. Alcuni Na+ e Cl- sono persi nell’urina, ma questa perdita è compensata dall’assorbimento attivo di sale disciolto dalle branchie. Nei crostacei marini, come le aragoste e i granchi, il rene funziona per regolare la composizione salina dell’emolinfa attraverso la modifica selettiva del contenuto salino dell’urina tubulare. In queste forme l’urina rimane isosmotica rispetto al sangue.
Sistema nervoso e sensoriale: I sistemi nervosi dei crostacei e degli anellidi hanno molto in comune, anche se quelli dei crostacei hanno più fusione di gangli (Figura 19-5). Il cervello è una coppia di gangli sovraesofagei che fornisce nervi agli occhi e a due paia di antenne. È unito da connettivi al ganglio subesofageo, una fusione di almeno cinque paia di gangli che forniscono nervi alla bocca, alle appendici, all’esofago e alle ghiandole antennali. Il doppio cordone nervoso ventrale ha un paio di gangli per ogni somite e nervi che servono le appendici, i muscoli e altre parti.
In aggiunta a questo sistema centrale, ci può essere un sistema nervoso simpatico associato al tratto digestivo.
I crostacei hanno organi di senso meglio sviluppati rispetto agli anellidi. I più grandi organi di senso dei gamberi sono gli occhi e gli statocisti. Ampiamente distribuiti sul corpo sono i peli tattili, delicate proiezioni di cuticola che sono particolarmente abbondanti sulle chele, sull’apparato boccale e sul telson. I sensi chimici del gusto e dell’olfatto si trovano in peli sulle antenne, sull’apparato boccale e in altri luoghi.
Una statocisti simile a una sacca, che si apre alla superficie tramite un poro dorsale, si trova sul segmento basale di ogni prima antenna dei gamberi. La statocisti contiene una cresta che porta peli sensoriali formati dal rivestimento chitinoso e granelli di sabbia che servono come statoliti. Ogni volta che l’animale cambia la sua posizione, i cambiamenti corrispondenti nella posizione dei grani sui peli sensoriali sono trasmessi come stimoli al cervello, e l’animale può regolarsi di conseguenza. Ogni muta (ecdisi) della cuticola comporta la perdita del rivestimento cuticolare della statocisti e con esso i granelli di sabbia. Nuovi granelli sono raccolti attraverso il poro dorsale dopo l’ecdisi.
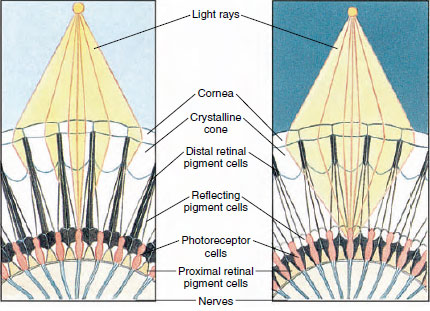
Figura 19-8
Parte dell’occhio composto di un artropode che mostra la migrazione del pigmento
negli ommatidi per la visione diurna e notturna. Cinque ommatidi sono rappresentati in
ogni diagramma. Di giorno ogni ommatidio è circondato da un collare scuro di
pigmento in modo che ogni ommatidio sia stimolato solo dai raggi di luce
che entrano nella sua cornea (visione a mosaico); di notte, il pigmento forma collari
incompleti e i raggi di luce possono diffondersi agli ommatidi adiacenti
(immagine continua, o sovrapposizione).
Gli occhi in molti crostacei sono composti, composti da molte unità di fotorecettori chiamate ommatidi (Figura 19-8). La superficie arrotondata di ogni occhio è coperta da un’area trasparente di cuticola, la cornea, che è divisa in molti piccoli quadrati o esagoni conosciuti come faccette. Queste faccette sono le estremità esterne degli ommatidi. Ogni ommatidio si comporta come un piccolo occhio e contiene diversi tipi di cellule disposte in modo colonnare (Figura 19-8). Le cellule del pigmento nero si trovano tra gli ommatidi adiacenti.
Il movimento del pigmento in un occhio composto artropode permette di adattarsi a diverse quantità di luce. Ci sono tre serie di cellule di pigmento in ogni ommatidio: retina distale, retina prossimale e riflettente; queste sono disposte in modo tale da formare un collare o un manicotto più o meno completo intorno ad ogni ommatidio. Per una forte luce o per l’adattamento al giorno, il pigmento retinico distale si muove verso l’interno e incontra il pigmento retinico prossimale che si muove verso l’esterno, in modo da formare un manicotto di pigmento completo attorno all’ommatidio (Figura 19-8). In questa condizione solo i raggi che colpiscono direttamente la cornea raggiungeranno le cellule fotorecettrici (retinulari), perché ogni ommatidio è schermato dagli altri. Così ogni ommatidio vedrà solo un’area limitata del campo visivo (un mosaico, o immagine di apposizione). In luce fioca i pigmenti distali e prossimali si separano in modo che i raggi di luce, con l’aiuto delle cellule pigmentarie riflettenti, abbiano la possibilità di diffondersi agli ommatidi adiacenti e di formare un’immagine continua, o di sovrapposizione. Questo secondo tipo di visione è meno preciso ma sfrutta al massimo la quantità limitata di luce ricevuta.
Riproduzione, cicli vitali e funzione endocrina
La maggior parte dei crostacei ha sessi separati, e ci sono varie specializzazioni per la copulazione tra i diversi gruppi. I cirripedi sono monoici ma generalmente praticano la fecondazione incrociata. In alcuni ostracodi i maschi sono scarsi e la riproduzione è solitamente partenogenetica. La maggior parte dei crostacei cova le uova in qualche modo: i branchiopodi e i cirripedi hanno speciali camere di cova, i copepodi hanno sacche di cova attaccate ai lati dell’addome (vedi Figura 19-19), e molti malacostraci portano uova e piccoli attaccati alle loro appendici addominali.
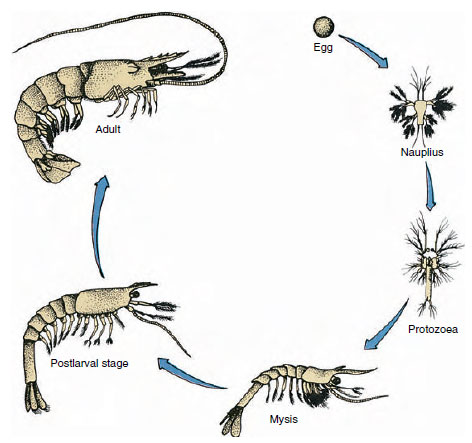
Figura 19-9
Ciclo vitale di un gambero del Golfo Penaeus. I peneidi si riproducono a profondità da 40 a 90 m. Le giovani forme larvali sono
planctoniche e si spostano verso la costa in acque con salinità inferiore per svilupparsi come giovani. I gamberi più vecchi ritornano in acque più profonde
offshore.
I pesci gatto hanno uno sviluppo diretto: non c’è una forma larvale. Un minuscolo giovane con la stessa forma dell’adulto e un set completo di appendici e somiti si schiude dall’uovo. Tuttavia, lo sviluppo è indiretto nella maggior parte dei crostacei, e una larva molto diversa dall’adulto nella struttura e nell’aspetto si schiude dall’uovo. Il passaggio dalla larva all’adulto è la metamorfosi. La larva primitiva e più diffusa nei crostacei è il nauplio (Figura 19-9 e 19-23). I nauplii hanno solo tre paia di appendici: le prime antenne uniramose, le antenne biramose e le mandibole biramose. Tutte funzionano come appendici per il nuoto in questo stadio.
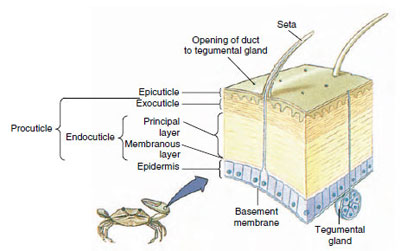
Figura 19-10
Struttura della cuticola del crostaceo.
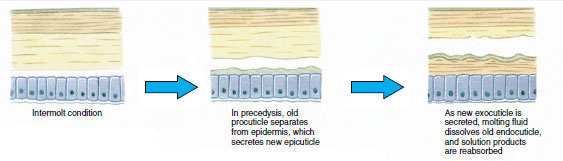
Lo sviluppo successivo può comportare un cambiamento graduale verso la forma del corpo adulto, e le appendici e i somiti vengono aggiunti attraverso una serie di mute, oppure l’assunzione della forma adulta può comportare cambiamenti più bruschi. Per esempio, la metamorfosi di un cirripede procede da un nauplio che nuota liberamente a una larva con un carapace bivalve chiamato cipride e infine a un adulto sessile con piastre calcaree: L’ecdisi (ek´duh-sis) (Gr. ekdyein, spogliarsi), o muta, è necessaria per aumentare le dimensioni del corpo perché l’esoscheletro non è vivente e non cresce con la crescita dell’animale. Gran parte del funzionamento di un crostaceo, compresa la riproduzione, il comportamento e molti processi metabolici, sono direttamente influenzati dalla fisiologia del ciclo di muta.
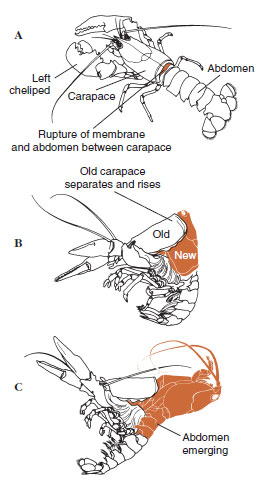
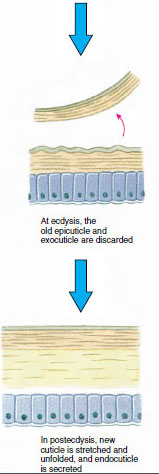
La cuticola, che è secreta dall’epidermide sottostante, ha diversi strati (Figura 19-10). Il più esterno è l’epicuticola, uno strato molto sottile di proteine impregnate di lipidi. La maggior parte della cuticola è costituita da diversi strati di procuticola: (1) esocuticola, che si trova appena sotto l’epicuticola e contiene proteine, sali di calcio e chitina; (2) endocuticola, che a sua volta è composta da (3) uno strato principale, che contiene più chitina e meno proteine ed è fortemente calcificato, e (4) uno strato membranoso non calcificato, uno strato relativamente sottile di chitina e proteine. Si separano dallo strato membranoso, secernono una nuova epicuticola e iniziano a secernere una nuova esocuticola (Figura 19-11). Gli enzimi vengono rilasciati nell’area sopra la nuova epicuticola. Questi enzimi iniziano a dissolvere la vecchia endocuticola, e i prodotti solubili vengono riassorbiti e immagazzinati all’interno del corpo del crostaceo. Alcuni sali di calcio sono immagazzinati come gastroliti (accrezioni minerali) nelle pareti dello stomaco. Alla fine, rimangono solo l’esocuticola e l’epicuticola della vecchia cuticola, sottoposta a nuova epicuticola e nuova esocuticola. L’animale inghiotte acqua, che assorbe attraverso l’intestino, e il suo volume sanguigno aumenta notevolmente. La pressione interna causa la spaccatura della cuticola, e l’animale si tira fuori dal suo vecchio esoscheletro (Figura 19-12). Seguono uno stiramento della nuova cuticola ancora morbida, la deposizione della nuova endocuticola, la rideposizione dei sali inorganici recuperati e di altri costituenti, e l’indurimento della nuova cuticola. Durante il periodo della muta, l’animale è indifeso e rimane nascosto.
 |
|
 |
 |
| Figura 19-12 Seguenza della muta nell’aragosta, Homarus americanus. A, la membrana tra il carapace e l’addome si rompe, e il carapace inizia una lenta elevazione. Questa fase può richiedere fino a 2 ore. B e C, la testa, il torace e infine l’addome si ritirano. Questo processo di solito non dura più di 15 minuti. Immediatamente dopo l’ecdisi, i chelipedi sono essiccati e il corpo è molto morbido. L’aragosta continua il rapido assorbimento di acqua in modo che entro 12 ore il corpo aumenta di circa il 20% in lunghezza e il 50% in peso. L’acqua del tessuto sarà sostituita da proteine nelle settimane successive. |
Figura 19-11 Secrezione di cuticole e riassorbimento in ecdisi. |
Quando un crostaceo è giovane, l’ecdisi deve avvenire frequentemente per permettere la crescita, e il ciclo di muta è relativamente breve. Quando l’animale si avvicina alla maturità, i periodi di interscambio diventano progressivamente più lunghi, e in alcune specie la muta cessa del tutto. Durante i periodi di intermittenza, l’aumento della massa dei tessuti si verifica quando il tessuto vivo sostituisce l’acqua.
Controllo ormonale del ciclo di ecdisi
Anche se l’ecdisi è controllata ormonalmente, il ciclo è spesso iniziato da uno stimolo ambientale percepito dal sistema nervoso centrale. Tali stimoli possono includere la temperatura, la lunghezza del giorno e l’umidità (nel caso dei granchi terrestri). Il segnale dal sistema nervoso centrale diminuisce la produzione di un ormone che inibisce la muta dall’organo X. L’organo X è un gruppo di cellule neurosecretrici nel midollo allungato del cervello. Nei gamberi e in altri decapodi, il midollo terminale si trova nel peduncolo oculare. L’ormone è trasportato negli assoni dell’organo X alla ghiandola del seno (che probabilmente non ha funzione ghiandolare), anch’essa nel peduncolo oculare, dove è rilasciato nell’emolinfa.
Un calo del livello dell’ormone che inibisce la muta promuove il rilascio di un ormone della muta dagli organi Y. Gli Yorgans si trovano sotto l’epidermide vicino ai muscoli adduttori delle mandibole, e sono omologo alle ghiandole protoraciche degli insetti, che producono l’ormone ecdysone. L’azione dell’ormone della muta consiste nell’avviare i processi che portano all’ecdisi (proecdisi). Una volta iniziato, il ciclo procede automaticamente senza ulteriori azioni di ormoni dagli organi X o Y.
Altre funzioni endocrine: Non solo la rimozione dei peduncoli oculari accelera la muta, ma oltre 100 anni fa è stato scoperto che i crostacei a cui sono stati rimossi i peduncoli oculari non possono più adattare la colorazione del corpo alle condizioni di fondo. Più di 50 anni fa si scoprì che il difetto era causato non dalla perdita della vista ma dalla perdita di ormoni negli occhi. Il colore del corpo dei crostacei è in gran parte il risultato di pigmenti in speciali cellule ramificate (cromatofori) nell’epidermide.
La concentrazione di granuli di pigmento nel centro delle cellule causa un effetto di schiarimento, e la dispersione del pigmento in tutte le cellule causa un effetto di scurimento. Il comportamento del pigmento è controllato da ormoni provenienti da cellule neurosecretrici nel peduncolo oculare, così come la migrazione del pigmento retinico per l’adattamento alla luce e al buio negli occhi (Figura 19-8).
Il rilascio di materiale neurosecretorio dagli organi pericardici nella parete del pericardio causa un aumento della frequenza e dell’ampiezza del battito cardiaco.
Ghiandole androgene, trovate per la prima volta in un anfipode (Orchestia, un comune saltatore da spiaggia), si trovano nei malacostracani maschi. A differenza della maggior parte degli altri organi endocrini nei crostacei, questi non sono organi neurosecretori. La loro secrezione stimola l’espressione delle caratteristiche sessuali maschili. I giovani malacostraci hanno ghiandole androgene rudimentali, ma nelle femmine queste ghiandole non si sviluppano. Se vengono impiantate artificialmente in una femmina, le sue ovaie si trasformano in testicoli e iniziano a produrre sperma, e le sue appendici iniziano ad assumere caratteristiche maschili alla successiva muta. Negli isopodi le ghiandole androgene si trovano nei testicoli; in tutti gli altri malacostraci si trovano tra i muscoli dei coxopodi delle ultime zampe toraciche e in parte attaccate vicino alle estremità dei vasa deferentia. Anche se le femmine non possiedono organi simili alle ghiandole androgene, le loro ovaie producono uno o due ormoni che influenzano le caratteristiche sessuali secondarie.
Ormoni che influenzano altri processi corporei nei crostacei possono essere presenti, e l’evidenza suggerisce che una sostanza neurosecretoria prodotta nell’occhio regola il livello di zucchero nel sangue.
Abitudini alimentari
Le abitudini alimentari e gli adattamenti all’alimentazione variano notevolmente tra i crostacei. Molte forme possono passare da un tipo di alimentazione all’altro a seconda dell’ambiente e della disponibilità di cibo, ma tutte usano lo stesso set fondamentale di apparato boccale. Mandibole e mascelle servono per ingerire il cibo; i maxillipedi trattengono e schiacciano il cibo. Nei predatori le zampe che camminano, in particolare i chelipedi, servono per catturare il cibo.
Molti crostacei, sia grandi che piccoli, sono predatori, e alcuni hanno interessanti adattamenti per uccidere le prede. Una forma simile al gambero, Lygiosquilla, ha su una delle sue zampe un dito specializzato che può essere disegnato in una scanalatura e rilasciato improvvisamente per perforare la preda di passaggio. I gamberi pistola (Alpheus spp.) hanno una chela enormemente allargata che può essere armata come il martello di una pistola e fatta scattare con una forza che stordisce la loro preda.
Il cibo dei mangiatori in sospensione varia dal plancton e detriti ai batteri. I predatori consumano larve, vermi, crostacei, lumache e pesci. Gli spazzini si nutrono di materia animale e vegetale morta. I mangiatori in sospensione, come i gamberetti fata, le pulci d’acqua e i cirripedi, usano le loro gambe, che portano una spessa frangia di setole, per creare correnti d’acqua che spazzano le particelle di cibo attraverso le setole. I gamberi di fango (Upogebia spp.) usano lunghe setae sulle loro prime due paia di appendici toraciche per filtrare il materiale alimentare dall’acqua fatta circolare attraverso la loro tana dai movimenti dei loro nuotatori.
I gamberi hanno uno stomaco in due parti (Figura 19-13). La prima parte contiene un mulino gastrico in cui il cibo, già strappato dalle mandibole, può essere ulteriormente macinato da tre denti calcarei in particelle abbastanza fini da passare attraverso un filtro setoso nella seconda parte; le particelle di cibo passano poi nell’intestino per la digestione chimica.
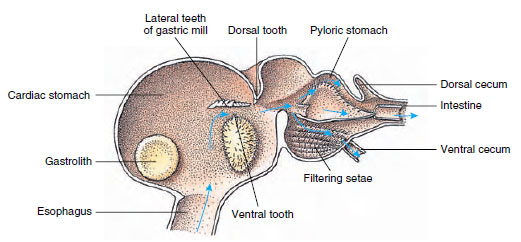
Figura 19-13
Stomaco malacostano che mostra il “mulino” gastrico e le direzioni dei movimenti del cibo. Il mulino ha creste chitinose, o
denti, per la masticazione, e setae per filtrare il cibo prima che passi nello stomaco pilorico.
Lascia un commento