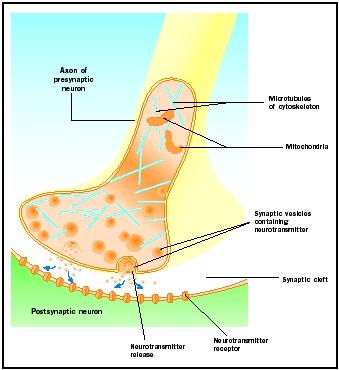
Neurotranszmitter felszabadulás
A szinaptikus csomókhoz érkező akciós potenciálok kiváltják a neurotranszmitter felszabadulását a szinaptikus hasadékba. A molekuláris mechanizmus nem teljesen tisztázott. Az akciós potenciál megérkezése és a neurotranszmitter felszabadulása között egy-két milliszekundumos “szinaptikus késleltetés” lép fel. Az akciós potenciálok megnyitják a kalciumcsatornákat a szinaptikus csomó membránjában, ami a kalciumionok befelé irányuló mozgását okozza. A kalciumionok kiváltják a neurotranszmitter felszabadulását a szinaptikus vezikulákból a szinaptikus hasadékba. Az exocitózis folyamata során a szinaptikus vezikulák összeolvadnak a preszinaptikus membránnal. A régi vezikulák membránjai a preszinaptikus membrán részévé válnak, az új vezikulák pedig a membrán egy szomszédos területéről csípődnek le. Ezek az új vezikulák később újonnan szintetizált vagy “újrahasznosított” neurotranszmitterekkel töltődnek fel .
A felszabadult neurotranszmitterek a szűk szinaptikus hasadékon keresztül diffundálnak. A posztszinaptikus membránon a neurotranszmitter molekulák az adott neurotranszmitterre specifikus felismerőhelyekkel rendelkező, membránhoz kötött receptormolekulákhoz kötődnek. A neurotranszmitter receptorhoz való kötődése az adott receptorra specifikus posztszinaptikus választ vált ki. Ezek a válaszok a receptor tulajdonságaitól függően lehetnek gerjesztőek vagy gátlóak. Ha a receptor stimulációja azt eredményezi, hogy a posztszinaptikus membrán elektromosan pozitívabbá válik (depolarizálódik), akkor ez egy gerjesztő posztszinaptikus potenciál (EPSP). Ha negatívabb (hiperpolarizált), akkor gátló posztszinaptikus potenciálról (IPSP) van szó. A gerjesztés és a gátlás a receptor és nem a neurotranszmitter tulajdonságaitól függ. A nátrium- vagy kalciumcsatornákhoz kapcsolt receptorok gerjesztő hatásúak és a posztszinaptikus membrán depolarizációját okozzák, míg a klorid- vagy káliumcsatornákhoz kapcsolt receptorok gátló hatásúak és a posztszinaptikus membrán hiperpolarizációját okozzák. Az ilyen ioncsatornákhoz kapcsolt receptorokat ionotróp receptoroknak nevezzük.
Más receptorok “második hírvivő” rendszerekhez kapcsolódnak, amelyek biokémiai reakciók sorozatát indítják el a posztszinaptikus sejtben. Ezek a metabotróp receptorok . A metabotróp receptorok sokféle
posztszinaptikus események. Ezek a szomszédos ioncsatornák közvetlen aktiválásától kezdve a receptorérzékenység megváltoztatásán át a specifikus hírvivő ribonukleinsavak (RNS-ek) átíródásáig, vagy akár specifikus gének aktiválásáig terjednek . A kémiai szinapszisok egy nagyon alkalmazkodó és rugalmas kommunikációs rendszer részét képezik. Ezek nem statikus anatómiai struktúrák rögzített tulajdonságokkal, hanem dinamikus struktúrák, amelyek a változó körülmények hatására képesek megváltoztatni molekuláris tulajdonságaikat.
Szó szerint több száz neurotranszmitter létezik. Néhányuk meglehetősen egyszerű vegyület, mint például az acetilkolin, a szerotonin, a katekolaminok (dopamin, noradrenalin és adrenalin) és számos aminosav . Sok közülük összetettebb, és a neuropeptid transzmitterek széles skálájához tartozik. A neurotranszmitterek a szinaptikus hasadékba való felszabadulás után mindaddig aktívak maradnak, amíg kémiailag meg nem változnak, vagy speciális szállítórendszerek által vissza nem kerülnek a szinaptikus csomóba, és újrahasznosításra nem kerülnek. A kolinerg szinapszisokban az acetilkolinészteráz jelen van a szinaptikus hasadékban. Ez az enzim a neurotranszmittert acetátra és kolinra hasítja, amelyek közül egyik sem aktív. A szerotonin és az adrenalin viszont a preszinaptikus terminálba kerül és újrahasznosul.
Vélemény, hozzászólás?