Subphylum Crustacea
A rákfélék általános természete
A rákfélék számos tekintetben különböznek a többi ízeltlábútól, de az egyetlen igazán megkülönböztető jellemzőjük az, hogy a rákfélék az egyetlen ízeltlábúak, amelyeknek két pár csápjuk van. A két pár csáp és egy pár állkapocs mellett a rákféléknek két pár állkapocs van a fejükön, majd egy pár függelék minden testszegmensen vagy szomitán. Egyes rákféléknél nem minden szomita visel függeléket. Az összes függelék, talán az első csáp kivételével, kezdetben birámás (két fő ágú), és legalábbis a mai kifejlett állatok néhány függeléke ezt az állapotot mutatja. A légzésre specializálódott szervek, ha vannak, kopoltyúként működnek.
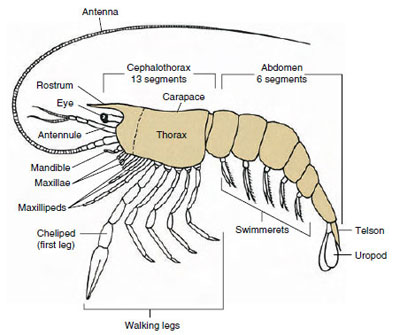
A legtöbb rákféle 16 és 20 szomita között van, de egyes formáknak 60 vagy több szomitájuk van. A nagyobb számú szomita primitív jellegzetesség. A származtatottabb állapot a kevesebb szegmentum és a fokozott tagmatizáció. A főbb tagmák a fej, a mellkas és a has, de ezek nem homológok az egész osztályban (vagy még egyes alosztályokon belül sem), mivel a szomiták különböző mértékben összeolvadnak, mint például a cephalothoraxban.
19-1. ábra
A Malacostraca archetipikus alaprajza. A két maxillae és a három
maxillipeda diagrammatikusan elkülönítettük, hogy szemléltessük a
általános tervet.
A rákfélék messze legnagyobb csoportja a Malacostraca osztály, amelybe a homárok, rákok, rákok, garnélák, parti szökevények, koca bogarak és még sokan mások tartoznak. Ezek a testszegmensek és tagmák meglepően állandó elrendezését mutatják, amit az osztály ősi tervének tartanak (19-1. ábra). Ez a tipikus testterv öt (embrionálisan hat) összenőtt szomitából álló fejet, nyolc szomitából álló mellkast és hat (néhány fajnál hét) szomitából álló hasat tartalmaz. Az elülső végén egy nem szegmentált rostrum, a hátsó végén pedig egy nem szegmentált telson található, amely az utolsó hasi szomittal és annak uropodáival sok formában egy farok legyezőt alkot.
Sok rákfélénél a fej háti kutikulája hátrafelé és az állat oldalai köré nyúlhat, hogy a mellkasi és hasi szomiták egy részét vagy egészét befedje vagy azokkal összeolvadjon. Ezt a burkolatot páncélnak nevezzük. Egyes csoportokban a páncél kagylóhéjszerű billentyűket alkot, amelyek a test nagy részét vagy az egész testet beborítják. A tízlábúaknál (beleértve a homárokat, garnélarákokat, rákokat és másokat) a páncél a teljes fejlábüreget fedi, de a hasüreget nem.
Forma és működés
Méretük és könnyű hozzáférhetőségük miatt a nagy rákokat, például a rákokat, más csoportoknál többet tanulmányozták. A bevezető laboratóriumi kurzusokon is gyakran tanulmányozzák őket. Ezért az alábbi megjegyzések közül sok kifejezetten a rákokra és rokonaikra vonatkozik.
Külső jellemzők
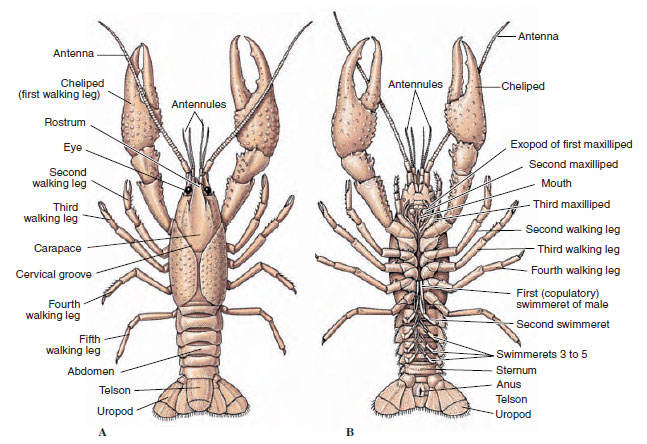
A rákok testét kitinből, fehérjéből és meszes anyagból álló, szekretált kutikula borítja. A nagyobb rákok keményebb, nehéz lemezei különösen sok meszet tartalmaznak. A kemény védőburkolat a szomiták közötti ízületeknél puha és vékony, ami lehetővé teszi a mozgás rugalmasságát. A páncél, ha van, a cephalothorax nagy részét vagy egészét fedi; a tízlábúaknál, például a rákoknál az összes fej- és mellkasi szegmentumot hátulról a páncél veszi körül. Minden egyes, a páncél által nem körülvett szomitát egy háti hámlemez, a tergum (19-2A. ábra) borít, és a szegmentális függelékek között egy ventrális keresztirányú sáv, a szegycsont helyezkedik el (19-2B. ábra). A hasüreg a telsonban végződik, amely nem tekinthető szomitának, és amelyen a végbélnyílás található. (A telson homológ lehet a gyűrűsgyűrűs pygidiummal.)
19-2. ábra
A rákok külső felépítése. A, Dorsalis nézet. B, Hasi nézet.
19-3. ábra
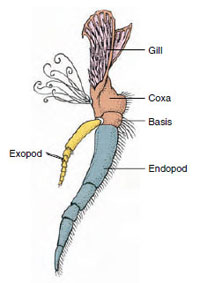
A birámás rákok
függelékének részei (a
rák harmadik maxillipedje).
A gonopórák elhelyezkedése nemenként és a rákok csoportjától függően változik. Lehetnek a függelékpáron vagy annak tövében, a test végső végén, vagy lábak nélküli szomitákon. A rákoknál a vasa deferentia nyílása a medián oldalon, az ötödik járóképes lábpár tövében, a petevezetékek nyílása pedig a harmadik pár tövében található. A nőstényeknél az ondócsatorna nyílása általában a negyedik és ötödik pár járóláb közötti középső vonalban található.
Függelékek: A Malacostraca (beleértve a rákokat is) és a Remipedia osztályok tagjainak jellemzően minden szomitán egy pár ízelt függelék található (19-3. ábra), bár a többi osztályban a hasi szomiták nem viselnek függeléket. A származtatott rákfélék, például a rákok függelékeiben jelentős specializáció figyelhető meg. Mindegyik azonban az alapvető, birámás terv változata, amit egy rákfüggelék, például a maxilliped (egy fejfüggelékké módosított mellkasi végtag) szemléltet (19-3. és 19-4. ábra). A bazális rész, vagy protopod, egy laterális exopodot és egy mediális endopodot hordoz. A protopod egy vagy két ízületből (basis és coxa) áll, míg az exopod és az endopod egytől több ízületig terjed. Néhány függelék, mint például a rákok járólábai, másodlagosan egyszárnyúvá váltak. A rákok végtagjain néha mediális vagy laterális nyúlványok fordulnak elő, amelyeket enditáknak, illetve exitáknak neveznek, és a protopodán lévő exitát epipodának nevezik. Az epipódok gyakran kopoltyúkká módosulnak. A 19-1. táblázat mutatja, hogy a különböző függelékek hogyan módosultak a birámiás tervből, hogy meghatározott funkcióknak megfeleljenek.
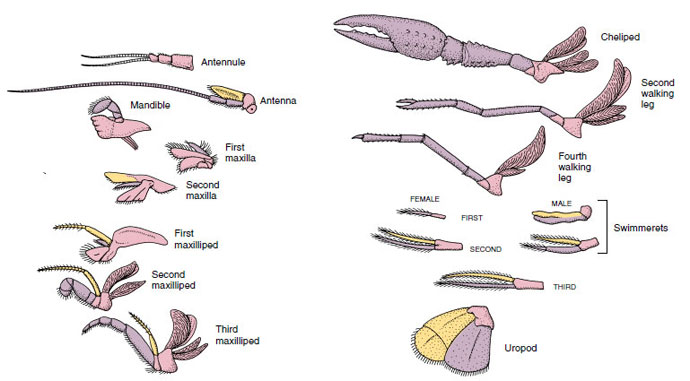
19-4. ábra
A rák függelékei, amelyeken látható, hogyan módosultak az alap birámiás tervből, ahogyan az egy
úszóhalban található. Protopod, rózsaszín; endopod, lila; exopod, sárga.
A hasonló alaprajzzal rendelkező és egy közös formából leszármazott szerkezeteket homológnak mondjuk, függetlenül attól, hogy azonos funkcióval rendelkeznek-e vagy sem. Mivel a specializált járólábak, a szájszervek, a csápok és az úszók mind egy közös birámatípusból fejlődtek ki, de különböző funkciók ellátására módosultak, mind homológok egymással, ezt az állapotot nevezzük soros homológiának. Eredetileg a végtagok mind nagyon hasonlóak voltak, de a szerkezeti módosulások evolúciója során egyes elágazások csökkentek, mások eltűntek, mások nagymértékben megváltoztak, és néhány új rész hozzáadódott. A rákok és szövetségeseik rendelkeznek a legfejlettebb sorozatos homológiával az állatvilágban, 17 különböző, de sorozatosan homológ végtagtípussal rendelkeznek (19-1. táblázat).
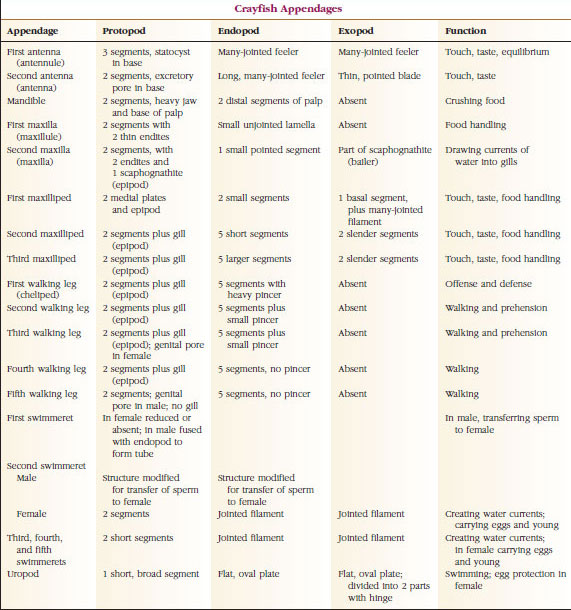
19. TÁBLÁZAT.1
Belső tulajdonságok
Az izom- és idegrendszer, valamint a mellkas és a has szegmentáció egyértelműen a gyűrűsgyíkszerű ősöktől örökölt metamerizmust mutatja, de más rendszerekben markáns módosulások vannak. A legtöbb változás a részek egy adott régióba való koncentrálódását, illetve a részek, mint például az intersepta, csökkenését vagy teljes elvesztését jelenti.
Hemocoel: Az ízeltlábúak fő testtere nem a coelom, hanem a vérrel teli hemocoel. A legtöbb ízeltlábú embrionális fejlődése során legalább néhány szomita mezodermájában csökevényes coelomikus üregek nyílnak. Ezek hamarosan eltűnnek, vagy folytonossá válnak a fejlődő mezodermális és ektodermális struktúrák és a sárgatest közötti térrel. Ez a tér lesz a hemocoel, és így nem béleli mezodermális peritoneum. A rákfélékben az egyetlen megmaradt coelomikus rekeszek a kiválasztó szervek végzsákjai és az ivarmirigyek körüli tér.
Muszkuláris rendszer: A legtöbb Crustacea testének jelentős részét csíkozott izmok alkotják. Az izmok általában antagonisztikus csoportokba rendeződnek: flexorok, amelyek egy testrészt a test felé húznak, és extenzorok, amelyek kifelé nyújtják azt. A rákok hasa erőteljes hajlítóizmokkal rendelkezik (19-5. ábra), amelyeket akkor használnak, amikor az állat hátrafelé úszik – ez a legjobb módja a menekülésnek. A gyomor mindkét oldalán erős izmok irányítják az állkapcsokat.
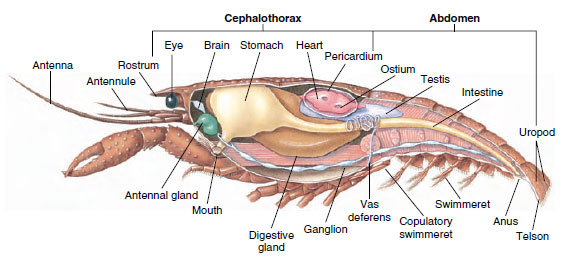
19-5. ábra
A hím rák belső felépítése.
Légzőrendszer: A légzőgázcsere a kisebb rákoknál a kutikula vékonyabb területein (például a lábakon) vagy az egész testen történik, és a specializált struktúrák hiányozhatnak. A nagyobb rákfélék kopoltyúkkal rendelkeznek, amelyek nagyon vékony hámréteggel rendelkező, finom, tollszerű nyúlványok. A tízlábúaknál a páncél oldalai zárják körül a kopoltyúüreget, amely elülső és hasi irányban nyitott (19-6. ábra). A kopoltyúk benyúlhatnak a mellhártya falából a kopoltyúüregbe, a mellkasi lábak és a test közötti csuklóból vagy a mellkasi bütykökből. Az utóbbi két típus jellemző a rákokra. A második felső állkapocs egy része, a “bailer” a kopoltyúfonalakon át, a lábak tövénél a kopoltyúüregbe, a lábak tövénél pedig a kopoltyúüregből az elülső részen húzza ki a vizet.
Keringési rendszer: A rákfélék és más ízeltlábúak keringési rendszere “nyitott” vagy lacunáris típusú. Ez azt jelenti, hogy nincsenek vénák, és nincs elkülönítve a vér és az interstitialis folyadék, mint a zárt rendszerű állatoknál. A hemolimpha (vér) az artériákon keresztül hagyja el a szívet, a hemocoeleken keresztül kering, és vénák helyett vénás szinuszokba vagy terekbe tér vissza, mielőtt újra belépne a szívbe. A gerincesekhez hasonlóan a gerincesek is zárt rendszerűek.
A háti szív a fő hajtószerv. Ez egy egykamrás, harántcsíkolt izomzatú zsák. A hemolimpha a környező szívburok szinuszból lép be a szívbe páros ostiumokon keresztül, olyan szelepekkel, amelyek megakadályozzák a szinuszba való visszaáramlást (19-6. ábra). A szívből a hemolimma egy vagy több artériába jut. Az artériákban lévő szelepek megakadályozzák a hemolimfa visszaáramlását. A kis artériák a szöveti szinuszokba ürülnek, amelyek viszont gyakran egy nagy szegycsonti szinuszba torkollnak (19-6. ábra).
Az afferens szinuszcsatornák onnan a hemolimfát a kopoltyúkhoz szállítják, ha vannak, oxigén- és szén-dioxid-csere céljából. A hemolimfa ezután az efferens csatornákon keresztül visszatér a szívburok szinuszába (19-6. ábra).
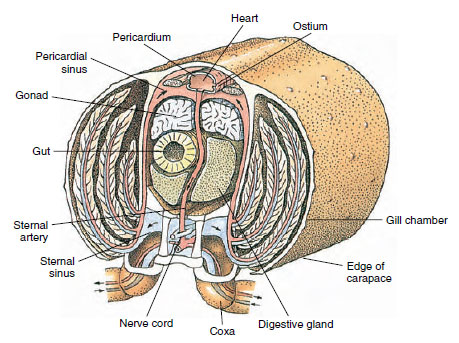
19-6. ábra
Diagrammatikus keresztmetszet a rák szív tájékán, amely a véráramlás irányát mutatja ebben a “nyitott”
vérrendszerben. A szív az artériákon keresztül pumpálja a vért a testszövetekbe, amelyek a szöveti szinuszokba ürülnek. A visszatérő
vér a szegycsonti szinuszba jut, majd a kopoltyúkon keresztül a gázcseréhez, és végül az efferens
csatornákon keresztül vissza a szívburok szinuszba. Figyeljük meg a vénák hiányát.
A hemolimpha az ízeltlábúaknál nagyrészt színtelen. Legalább kétféle ameboid sejtet tartalmaz. A hemocianin, egy réztartalmú légzőszervi pigment, vagy a hemoglobin, egy vastartalmú pigment, oldatban hordozható. A hemolimfának megvan az alvadási tulajdonsága, ami megakadályozza az elvesztését kisebb sérülések esetén. Egyes ameboid sejtek trombinszerű alvadékot bocsátanak ki, amely elősegíti az alvadást.
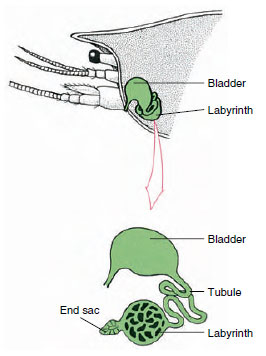
19-7. ábra
A rákok antennamirigyének (zöld mirigyének)
Sémája. (Természetes helyzetben a szerv sokkal
behajtott.) Egyes rákoknál hiányzik a labirintus,
és a kiválasztócső (nephridialis csatorna)
sok tekervényes cső.
Kiválasztórendszer: A kifejlett rákok kiválasztó szervei a fej ventrális részén, a nyelőcső előtt elhelyezkedő csöves szerkezetek párja (19-5. ábra). Attól függően, hogy az antennák vagy a második állkapocs tövében nyílnak-e, antennamirigyeknek vagy maxilláris mirigyeknek nevezik őket. Néhány kifejlett rákféle mindkettővel rendelkezik. A tízlábúak kiválasztó szervei az antennamirigyek, amelyeket ebben a csoportban zöldmirigyeknek is neveznek. A rákféléknek nincsenek malpighian tubulusai, a pókok és rovarok kiválasztó szervei.
Az embrionális coelomikus rekeszből származó antennamirigy végzsákja egy kis hólyagocskából (saccula) és egy szivacsos masszából, az úgynevezett labirintusból áll. A labirintus egy kiválasztó tubuluson keresztül egy dorzális hólyaghoz csatlakozik, amely a bazális antennaszegmens ventrális felületén lévő póruson keresztül nyílik a külvilág felé (19-7. ábra). A hemocoelben lévő hidrosztatikus nyomás biztosítja a folyadéknak a végzsákba történő szűrését. Ahogy a szűrlet áthalad a kiválasztó tubuluson és a hólyagon, a sók, aminosavak, glükóz és némi víz reszorpciójával módosul, és végül vizeletként ürül ki.
A nitrogéntartalmú hulladékok (főként ammónia) kiválasztása a kutikula vékony területein, különösen a kopoltyúkon keresztül történő diffúzióval történik, és az úgynevezett kiválasztószervek elsősorban a testnedvek ionos és ozmotikus összetételének szabályozására szolgálnak. Az édesvízi rákokat, például a rákokat, folyamatosan fenyegeti a vérük túlhígulása a kopoltyúkon és más vízáteresztő felületeken keresztül diffundáló vízzel. A zöld mirigyek híg, alacsony sótartalmú vizeletet képezve hatékony “árvízvédelmi” eszközként működnek. A vizelettel némi Na+ és Cl- elvész, de ezt a veszteséget az oldott sónak a kopoltyúk általi aktív felszívódása kompenzálja. A tengeri rákoknál, például a homároknál és a rákoknál a vese a hemolimfa sóösszetételét a csöves vizelet sótartalmának szelektív módosításával szabályozza. Ezekben a formákban a vizelet a vérrel szemben izoszmotikus marad.
Ideg- és érzékszervi rendszerek: A rákfélék és a gyűrűsállatok idegrendszere sok közös vonást mutat, bár a rákfélék idegrendszerében több ganglion fúziója van (19-5. ábra). Az agy egy pár szupraesophageális ganglion, amely a szem és a két pár csáp idegeit látja el. Ehhez csatlakozik a suboesophagealis ganglion, amely legalább öt ganglionpár fúziója, amelyek a száj, a függelékek, a nyelőcső és az antennamirigyek idegeit látják el. A kettős ventrális idegszálban minden szomita számára egy pár ganglion található, és a függelékeket, izmokat és más részeket ellátó idegek.
A központi rendszeren kívül az emésztőrendszerhez szimpatikus idegrendszer is kapcsolódhat.
A rákfélék érzékszervei jobban fejlettek, mint a gyűrűsgyíkoké. A rákok legnagyobb érzékszervei a szemek és a statociszták. Széles körben eloszlanak a testen a tapintószőrök, a kutikula finom nyúlványai, amelyek különösen az állkapcsokon, a szájszervecskéken és a telsonon fordulnak elő. Az ízlelés és a szaglás kémiai érzékszervei az antennákon, a szájszerveken és más helyeken találhatóak szőrökben.
A rákok minden első antennájának bazális szegmensén egy-egy zsákszerű statociszta található, amely egy dorzális póruson keresztül nyílik a felszínre. A statociszta egy gerincet tartalmaz, amely a kitinszerű bélésből és homokszemcsékből kialakult érzékszőröket hordoz, amelyek statolitként szolgálnak. Amikor az állat megváltoztatja a helyzetét, az érzékszőrökön lévő szemcsék helyzetének megfelelő változásai ingerületként jutnak el az agyba, és az állat ennek megfelelően tud alkalmazkodni. A hám minden egyes vedlése (ekdízis) a statociszta hámbélésének és vele együtt a homokszemcséknek az elvesztésével jár. Az ekdízis után a dorzális póruson keresztül új szemcséket vesznek fel.
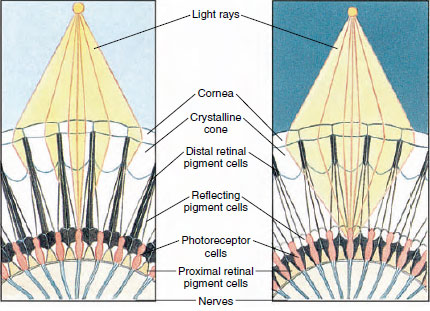
19-8. ábra
Az ízeltlábúak összetett szemének egy része, amely a pigment
vándorlását mutatja az ommatidiákban a nappali és éjszakai látáshoz. Öt ommatídiumot ábrázolnak
minden ábrán. Nappal minden egyes ommatídiumot sötét
pigmentgallér vesz körül, így minden egyes ommatídiumot csak a saját szaruhártyáján bejutó
fénysugarak stimulálnak (mozaikos látás); éjjel a pigment
nem teljes gallért alkot, és a fénysugarak átterjedhetnek a szomszédos ommatídiumokra
(folyamatos, vagy szuperpozíciós kép).
A szem sok rákfélében összetett, sok fotoreceptor egységből, úgynevezett ommatídiumokból áll (19-8. ábra). Minden szem lekerekített felületét egy áttetsző hámterület, a szaruhártya borítja, amely sok kis négyzetre vagy hatszögre, úgynevezett facettára van osztva. Ezek a facetták az ommatídiumok külső végei. Minden egyes ommatídium úgy viselkedik, mint egy apró szem, és többféle, oszlopszerűen elrendezett sejtet tartalmaz (19-8. ábra). A szomszédos ommatídiumok között fekete pigmentsejtek találhatók.
A pigment mozgása az ízeltlábúak összetett szemében lehetővé teszi a különböző fénymennyiségekhez való alkalmazkodást. Minden ommatídiumban három pigmentsejtcsoport található: distalis retinális, proximális retinális és fényvisszaverő; ezek úgy vannak elrendezve, hogy többé-kevésbé teljes gallért vagy hüvelyt alkothatnak minden ommatídium körül. Erős fény vagy nappali adaptáció esetén a distalis retinális pigment befelé mozog, és találkozik a kifelé mozgó proximális retinális pigmenttel, így egy teljes pigmenthüvely alakul ki az ommatidium körül (19-8. ábra). Ebben az állapotban csak a szaruhártyát közvetlenül érő sugarak érik el a fotoreceptor (retinuláris) sejteket, mivel minden egyes ommatidium le van árnyékolva a többitől. Így minden ommatidium a látómezőnek csak egy korlátozott területét látja (mozaikos vagy appozíciós kép). Gyenge fényben a disztális és proximális pigmentek szétválnak, így a fénysugaraknak a fényvisszaverő pigmentsejtek segítségével esélyük van arra, hogy a szomszédos ommatídiumok felé terjedjenek, és folyamatos vagy szuperpozíciós képet alkossanak. Ez a második típusú látás kevésbé pontos, de maximálisan kihasználja a befogadott korlátozott fénymennyiséget.
Reprodukció, életciklusok és endokrin működés
A legtöbb rákfélének különálló neme van, és a különböző csoportok között a kopulációra különböző specializációk léteznek. A pajzstetvek egylakiak, de általában gyakorolják a keresztmegtermékenyítést. Egyes ostracodáknál a hímek ritkák, és a szaporodás általában parthenogenetikus. A legtöbb rákféle valamilyen módon költözteti a tojásait: a branchiopodák és a pajorok speciális költőkamrákkal rendelkeznek, a kopárhátúak a hasüreg oldalára erősített költőzsákokkal rendelkeznek (lásd a 19-19. ábrát), és sok malacostracana a hasi függelékükre erősítve hordja a tojásokat és a kicsinyeket.
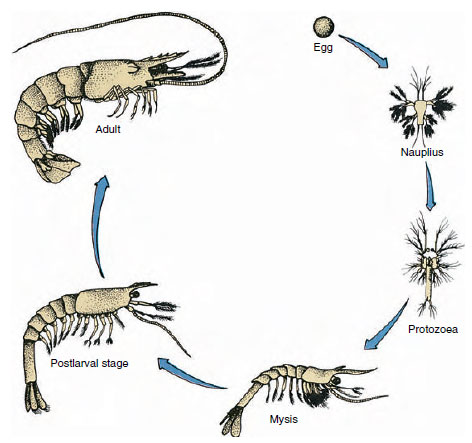
19-9. ábra
A Penaeus öbölgarnéla életciklusa. A penaeidák 40-90 m mélységben ívnak. A fiatal lárvaformák
planktonikusak, és a parton alacsonyabb sótartalmú vízbe vándorolnak, hogy ivadékként fejlődjenek. Az idősebb garnélarákok visszatérnek a mélyebb vízbe
a parttól távolabbra.
A rákok fejlődése közvetlen: nincs lárvaforma. Az ikrából egy apró, a kifejlett egyeddel azonos alakú, a függelékek és a szomiták teljes készletével rendelkező ivadék kel ki. A rákfélék többségénél azonban a fejlődés közvetett, és a tojásból egy olyan lárva kel ki, amely felépítésében és megjelenésében teljesen eltér a kifejlett egyedtől. A lárvából végül felnőtté válás a metamorfózis. A rákfélékben a legprimitívebb és leggyakrabban előforduló lárva a nauplius (19-9. és 19-23. ábra). A naupliák csak három pár függeléket viselnek: uniramikus első antennák, biramikus antennák és biramikus mandibulák. Ebben a stádiumban mindegyik úszó függelékként funkcionál.
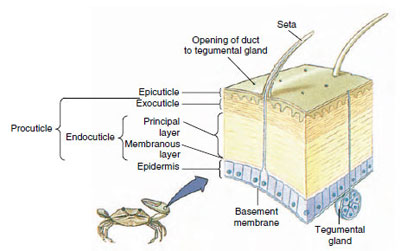
19-10. ábra
A rákfélék kutikulájának felépítése.
A későbbi fejlődés magában foglalhatja a kifejlett testforma fokozatos átalakulását, és a függelékek és a szomiták moltsorozaton keresztül kerülnek hozzá, vagy a kifejlett forma átvétele hirtelenebb változásokkal járhat. Például a pajzstetvek metamorfózisa a szabadon úszó naupliusból a kétkagylós páncéllal rendelkező lárvává, ún. cypridává, és végül a mészlemezekkel rendelkező, ülő felnőtt egyeddé válik.
Ecdysis: Az ekdízis (ek´duh-sis) (gr. ekdyein, levetkőzni) vagy vedlés szükséges a test méretének növekedéséhez, mivel a külső váz nem élő, és nem növekszik az állat növekedésével. A rákfélék működésének nagy részét, beleértve a szaporodást, a viselkedést és számos anyagcsere-folyamatot, közvetlenül befolyásolja a vedlési ciklus fiziológiája.
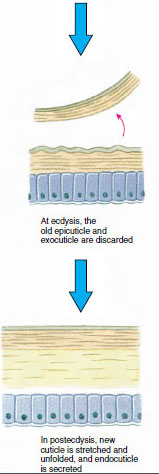
A hám, amelyet az alatta lévő epidermisz választ ki, több rétegből áll (19-10. ábra). A legkülső az epikutikula, egy nagyon vékony, lipidekkel impregnált fehérjeréteg. A kutikula nagy részét a prokutikula több rétege alkotja: (1) az exocuticle, amely közvetlenül az epicuticle alatt található, és fehérjét, kalciumsókat és kitint tartalmaz; (2) az endocuticle, amely maga is (3) egy fő rétegből áll, amely több kitint és kevesebb fehérjét tartalmaz, és erősen meszesedik, valamint (4) egy nem meszesedett hártyarétegből, egy viszonylag vékony kitin- és fehérjerétegből.
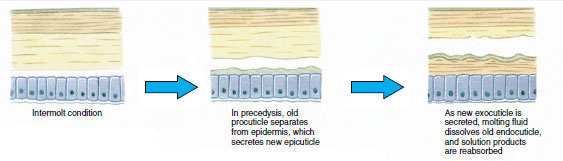
Mivel a tényleges ekdysis előtt az epidermális sejtek jelentősen megnagyobbodnak. Leválnak a hártyás rétegtől, új epikutikulumot választanak ki, és új exocutikulum kiválasztásába kezdenek (19-11. ábra). Az új epikutikulum feletti területre enzimek szabadulnak fel. Ezek az enzimek elkezdik a régi endocutikula feloldását, az oldódó termékek pedig felszívódnak és a rák testében tárolódnak. A kalciumsók egy része gasztrolitokként (ásványi lerakódások) tárolódik a gyomor falában. Végül a régi kutikulából csak az exocuticle és az epicuticle marad meg, amelyek alatt új epicuticle és új exocuticle található. Az állat vizet nyel, amelyet a bélrendszerén keresztül vesz fel, és a vértérfogata nagymértékben megnő. A belső nyomás hatására a kutikula felhasad, és az állat kihúzza magát a régi exoskeletonjából (19-12. ábra). Ezután a még mindig puha új kutikula megnyúlása, az új endokutikula lerakódása, a megmentett szervetlen sók és egyéb alkotóelemek újbóli lerakódása, valamint az új kutikula megkeményedése következik. A vedlés ideje alatt az állat védtelen, és rejtve marad.
 |
|
 |
 |
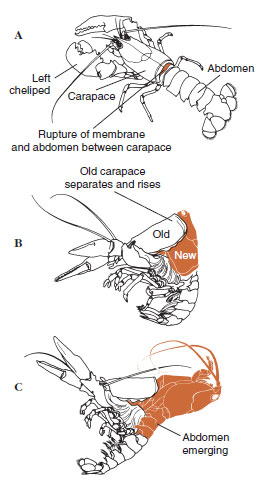
| 19-12. ábra A vedlés sorrendje a Homarus americanus homárban. A, A páncél és a has közötti hártya megreped, és a páncél lassan emelkedni kezd. Ez a lépés akár 2 órát is igénybe vehet. B és C, A fej, a mellkas és végül a has visszahúzódik. Ez a folyamat általában nem tart tovább 15 percnél. Közvetlenül az ekdízis után az állkapcsok kiszáradnak és a test nagyon puha. A homár folytatja a gyors vízfelvételt, így 12 óra alatt a test hossza kb. 20%-kal, tömege pedig 50%-kal nő. A szöveti vizet a következő hetekben fehérjével pótolják. |
19-11. ábra Hámszelválasztás és felszívódás az ekdízis során. |
Amikor a rák fiatal, az ekdízisnek gyakran kell bekövetkeznie, hogy növekedni tudjon, és a vedlési ciklus viszonylag rövid. Ahogy az állat közeledik az ivarérettséghez, a vedlés közötti időszakok fokozatosan hosszabbak lesznek, és egyes fajoknál a vedlés teljesen megszűnik. A két vedlés közötti időszakok alatt a szövetek tömege növekszik, mivel az élő szövetek helyettesítik a vizet.
Az ekdízisciklus hormonális irányítása
Bár az ekdízis hormonálisan szabályozott, a ciklust gyakran a központi idegrendszer által érzékelt környezeti inger indítja el. Ilyen ingerek lehetnek a hőmérséklet, a nap hossza és a páratartalom (szárazföldi rákok esetében). A központi idegrendszerből érkező jel hatására az X-szerv csökkenti az ívást gátló hormon termelését. Az X-szerv az agy medulla terminalisában található neuroszekréciós sejtek csoportja. A rákoknál és más tízlábúaknál a medulla terminalis a szemszárban található. A hormon az X-szerv axonjain keresztül jut el a sinus mirigyhez (amely maga valószínűleg nem mirigyes működésű), szintén a szemszárban, ahol a hemolimfába szabadul fel.
A molyhormon szintjének csökkenése elősegíti egy molyhormon felszabadulását az Y-szervekből. Az Yorgansok az epidermisz alatt, az állkapcsok adductor izmainak közelében találhatók, és homol-ogusak a rovarok prothoracalis mirigyeivel, amelyek az ecdysone hormont termelik. A vedlési hormon hatása az ecdysishez (proecdysis) vezető folyamatok elindítása. A ciklus a beindulás után automatikusan, az X- vagy Y-szervekből származó hormonok további hatása nélkül folytatódik.
Egyéb endokrin funkciók: Nemcsak a szemszárak eltávolítása gyorsítja fel a vedlést, hanem több mint 100 évvel ezelőtt azt is megállapították, hogy azok a rákok, amelyeknek a szemszárát eltávolították, már nem tudják a testük színét a háttérkörülményekhez igazítani. Több mint 50 évvel ezelőtt felfedezték, hogy a hibát nem a látás elvesztése, hanem a szemszárakban lévő hormonok elvesztése okozza. A rákok testszíne nagyrészt az epidermisz speciális, elágazó sejtjeiben (kromatofórák) található pigmenteknek köszönhető.
A pigmentszemcsék koncentrációja a sejtek közepén világosodást, a pigment szétszóródása a sejtekben pedig sötétedést okoz. A pigment viselkedését a szemhéjban lévő neuroszekréciós sejtekből származó hormonok irányítják, akárcsak a retinapigment vándorlását a szem világos és sötét adaptációjához (19-8. ábra).
A neuroszekréciós anyag felszabadulása a szívburok falában lévő perikardiális szervekből a szívverés ütemének és amplitúdójának növekedését okozza.
A hím malacostracáknál előfordulnak androgén mirigyek, amelyeket először egy kétéltűben (Orchestia, egy közönséges strandhopper) találtak. A rákfélék legtöbb más endokrin szervétől eltérően ezek nem neuroszekréciós szervek. Szekréciójuk serkenti a hím nemi jellegek kifejeződését. A fiatal malacostracánoknak csökevényes androgén mirigyei vannak, de a nőstényeknél ezek a mirigyek nem fejlődnek ki. Ha mesterségesen beültetjük őket egy nősténybe, a petefészkei herékké alakulnak át, és spermiumot kezdenek termelni, a következő vedléskor pedig a függelékei hímnemű tulajdonságokat vesznek fel. Az izopodáknál az androgén mirigyek a herékben találhatók; minden más malacostracánnál az utolsó mellkasi lábak coxopodiumainak izmai között találhatók, és részben a vasa deferentia végeihez kapcsolódnak. Bár a nőstények nem rendelkeznek az androgén mirigyekhez hasonló szervekkel, petefészkeik egy vagy két olyan hormont termelnek, amelyek befolyásolják a másodlagos nemi jellegeket.
A Crustaceáknál más testi folyamatokat befolyásoló hormonok is jelen lehetnek, és bizonyítékok utalnak arra, hogy a szemhéjban termelődő neuroszekréciós anyag szabályozza a vércukorszintet.
Táplálkozási szokások
A táplálkozási szokások és a táplálkozáshoz való alkalmazkodás nagyon eltérő a rákfélék között. Sok forma a környezet és a táplálék elérhetőségének függvényében képes egyik táplálkozási módról a másikra váltani, de mindegyik ugyanazt az alapvető szájszervkészletet használja. Az állkapcsok és a felső állkapcsok a táplálék felvételére szolgálnak; a felső állkapcsok a táplálékot megtartják és összezúzzák. A ragadozóknál a járólábak, különösen az állkapocslábak a táplálék elfogására szolgálnak.
Néhány rákféle, kicsi és nagy egyaránt, ragadozó, és némelyiküknek érdekes alkalmazkodása van a zsákmány megölésére. Az egyik rákszerű formának, a Lygiosquilla-nak az egyik járólábán van egy speciális ujj, amelyet be lehet húzni egy barázdába, és hirtelen el lehet engedni, hogy átszúrja az elhaladó zsákmányt. A pisztolyos garnéláknak (Alpheus spp.) egy hatalmasra megnagyobbodott csélájuk van, amelyet úgy lehet felhúzni, mint egy pisztoly kalapácsát, és olyan erővel pattintani, hogy elkábítja a zsákmányt.
A szuszpenziós táplálkozók tápláléka a planktontól a törmeléken át a baktériumokig terjed. A ragadozók lárvákat, férgeket, rákokat, csigákat és halakat fogyasztanak. A dögevők elpusztult állati és növényi anyagokat fogyasztanak. A szuszpenziós táplálkozók, mint például a tündérgarnélák, a vízibolhák és a pajzstetvek, lábukkal, amelyeken vastag szarucsomók vannak, vízáramlatokat hoznak létre, amelyek a táplálékrészecskéket a szarucsomókon keresztül sodorják. Az iszapgarnélák (Upogebia spp.) a mellkasi függelékeik első két párján lévő hosszú szőrszálakat arra használják, hogy az úszóhártyájuk mozgatásával kiszűrjék a táplálékanyagot az odújukban keringő vízből.
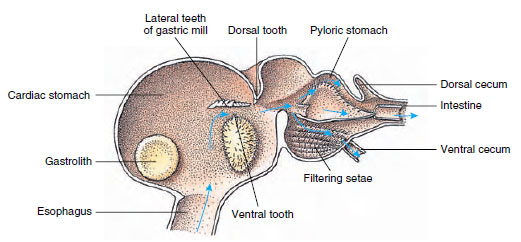
A rákoknak kétrészes gyomruk van (19-13. ábra). Az első rész egy gyomormalmot tartalmaz, amelyben az állkapcsok által már széttépett táplálékot három mészfog segítségével tovább őrölhetik olyan finomságú részecskékké, amelyek a második részben lévő szetoszűrőn átjutnak; a táplálékrészecskék ezután a bélbe jutnak kémiai emésztésre.
19-13. ábra
Malacostracan gyomor a gyomor “malmát” és a táplálékmozgások irányát mutatja. A malomnak kitines bordái vagy
fogai vannak a rágáshoz, és szőrszálai a táplálék szűréséhez, mielőtt az a pylorusos gyomorba kerülne.
Vélemény, hozzászólás?