Podtřída Crustacea
Obecná povaha korýšů
Korýši se od ostatních členovců liší mnoha způsoby, ale jediným skutečně charakteristickým znakem je, že korýši jsou jediní členovci se dvěma páry tykadel. Kromě dvou párů tykadel a páru čelistí mají korýši dva páry horních čelistí na hlavě a dále pár přívěsků na každém tělním článku nebo somitu. U některých korýšů nejsou všechny somity opatřeny přívěsky. Všechny přívěsky, snad s výjimkou prvních tykadel, jsou primitivně biramózní (dvě hlavní větve) a alespoň některé přívěsky současných dospělců vykazují tento stav. Orgány specializované na dýchání, pokud jsou přítomny, fungují jako žábry.
Většina korýšů má 16 až 20 somitů, ale některé formy mají 60 i více somitů. Větší počet somitů je primitivním znakem. Odvozenějším stavem je mít méně segmentů a zvýšenou tagmatizaci. Hlavními tagmaty jsou hlava, hrudník a břicho, které však nejsou v celé třídě (a dokonce ani v rámci některých podtříd) homologické, protože dochází k různému stupni splynutí somitů, například jako u hlavohrudi.
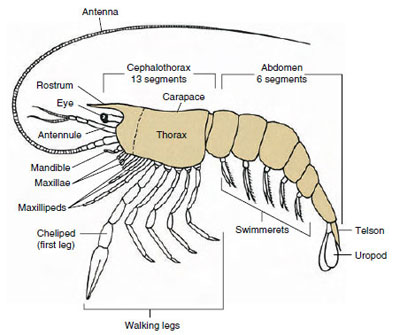
Obrázek 19-1
Archetypický plán Malacostraca. Dvě čelisti a tři
maxilipidy byly schematicky odděleny pro znázornění
obecného plánu.
Daleko největší skupinou korýšů je třída Malacostraca, do které patří humři, krabi, krevety, plážoví skákavky, rozsivky a mnoho dalších. Ti vykazují překvapivě konstantní uspořádání tělních segmentů a tagmat, které je považováno za ancestrální plán třídy (obr. 19-1). Tento typický tělní plán má hlavu z pěti (embryonálně ze šesti) srostlých somitů, hrudník z osmi somitů a břicho ze šesti somitů (u několika druhů ze sedmi). Na předním konci je nesegmentované rostrum a na zadním konci je nesegmentovaný telson, který s posledním břišním somitem a jeho uropody tvoří u mnoha forem ocasní vějíř.
U mnoha korýšů se může hřbetní kutikula hlavy rozšiřovat dozadu a kolem boků živočicha a zakrývat některé nebo všechny hrudní a břišní somity nebo s nimi splývat. Tento kryt se nazývá karapax. U některých skupin tvoří karapax škeblovité chlopně, které pokrývají většinu nebo celé tělo. U dekapodů (včetně humrů, krevet, krabů a dalších) pokrývá karapax celou hlavohruď, ale ne břicho.
Tvar a funkce
Velcí korýši, jako jsou raci, jsou díky své velikosti a snadné dostupnosti studováni více než jiné skupiny. Jsou také běžně studováni v úvodních laboratorních kurzech. Proto se mnohé z následujících poznámek týkají právě raků a jejich příbuzných.
Vnější znaky
Těla korýšů jsou pokryta vylučovanou kutikulou složenou z chitinu, bílkovin a vápenatých látek. Tvrdší a těžší desky větších korýšů jsou obzvláště bohaté na vápenaté usazeniny. Tvrdý ochranný povlak je na spojích mezi somity měkký a tenký, což umožňuje pružnost pohybu. Karapax, pokud je přítomen, pokrývá většinu nebo celou hlavohruď; u dekapodů, jako jsou raci, jsou všechny hlavové a hrudní segmenty hřbetně uzavřeny karapaxem. Každý somit, který není uzavřen karapaxem, je kryt hřbetní kutikulární destičkou neboli tergem (obrázek 19-2A) a mezi segmentálními přídavky leží břišní příčka, sternum (obrázek 19-2B). Břicho je zakončeno telsonem, který se nepovažuje za somit a nese řitní otvor. (Telson může být homologický s pygidiem annelidů.)
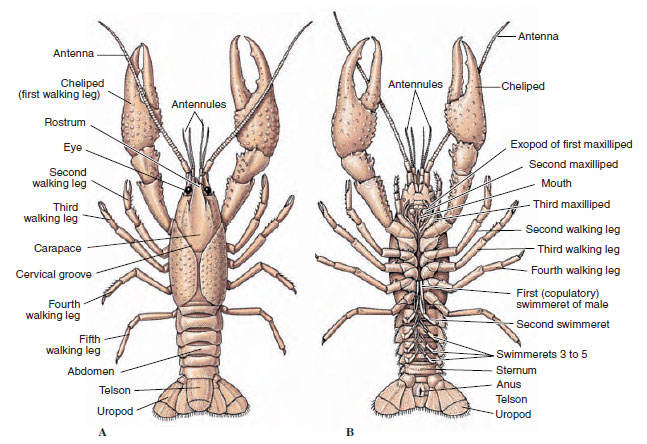
Obrázek 19-2
Vnější stavba raků. A, hřbetní pohled. B, Ventrální pohled.
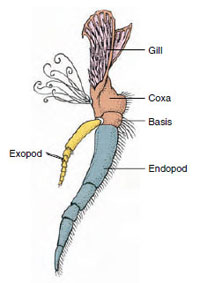
Obrázek 19-3
Části biramózního korýšího
apendixu (třetí maxiliped
raka).
Poloha gonopor se liší podle pohlaví a skupiny korýšů. Mohou být na bázi nebo na bázi páru přívěsků, na terminálním konci těla nebo na somitech bez nohou. U raků jsou otvory vasa deferentia na střední straně na bázi pátého páru kráčivých nohou a otvory vejcovodů na bázi třetího páru. U samic se otvor do semenné schránky obvykle nachází ve střední čáře mezi čtvrtým a pátým párem kráčivých nohou.
Přídatné orgány: Zástupci tříd Malacostraca (včetně raků) a Remipedia mají obvykle na každém somitu pár kloubních přívěsků (obrázek 19-3), ačkoli břišní somity ostatních tříd přívěsky nenesou. U odvozených korýšů, jako jsou raci, je patrná značná specializace výrůstků. Všechny jsou však variacemi základního, biramózního plánu, který ilustrují přídavky raků, jako je maxilliped (hrudní končetina modifikovaná na přídavek hlavy) (obrázky 19-3 a 19-4). Základní část neboli protopod nese laterální exopod a mediální endopod. Protopod se skládá z jednoho nebo dvou kloubů (base a coxa), zatímco exopod a endopod mají po jednom až několika kloubech. Některé končetiny, např. kráčivé nohy raků, jsou druhotně jednovrstevné. Na končetinách korýšů se někdy vyskytují mediální nebo laterální výběžky, které se nazývají endity, resp. exity, a exit na protopodu se nazývá epipod. Epipody jsou často modifikovány jako žábry. Tabulka 19-1 ukazuje, jak se různé výrůstky modifikovaly z biramusového plánu, aby vyhovovaly specifickým funkcím.
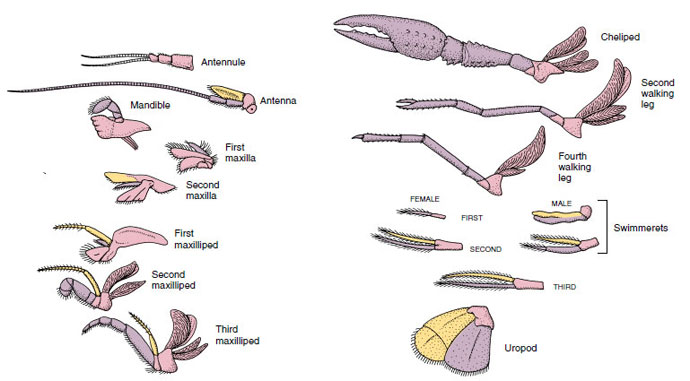
Obrázek 19-4
Přídavky raků ukazující, jak se modifikovaly ze základního biramózního plánu, jak je najdeme u
plavce. Protopod, růžový; endopod, fialový; exopod, žlutý.
O útvarech, které mají podobný základní plán a vznikly ze společné formy, se říká, že jsou homologické, ať už mají stejnou funkci, nebo ne. Vzhledem k tomu, že specializované kráčivé nohy, ústa, chelipody a plováky se vyvinuly ze společného biramusového typu, ale modifikovaly se tak, aby plnily různé funkce, jsou všechny navzájem homologické, což je stav známý jako sériová homologie. Primitivně byly všechny končetiny velmi podobné, ale během evoluce strukturálních modifikací se některé větve zredukovaly, některé ztratily, některé výrazně pozměnily a některé nové části přibyly. Raci a jejich příbuzní mají nejpropracovanější sériovou homologii v živočišné říši, mají 17 odlišných, ale sériově homologických typů končetin (tabulka 19-1).
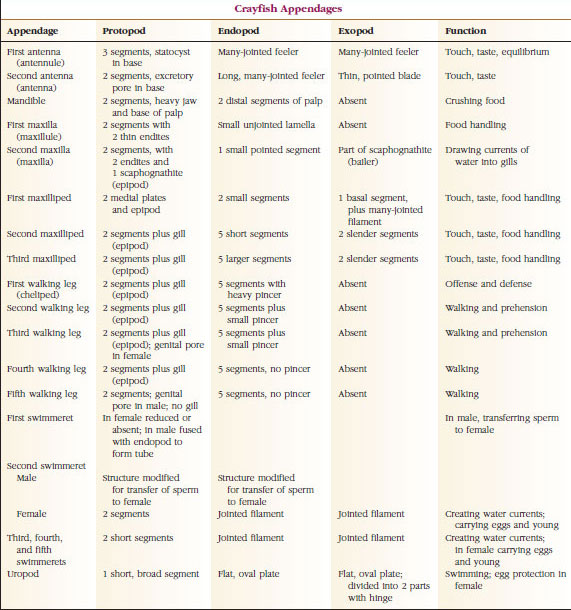
TABULKA 19. Sériová homologie končetin.1
Vnitřní znaky
Svalová a nervová soustava a segmentace v oblasti hrudníku a břicha zřetelně vykazují metamerismus zděděný po předcích podobných annelidům, ale v ostatních soustavách jsou výrazné modifikace. Většina změn zahrnuje soustředění částí v určité oblasti nebo naopak redukci či úplnou ztrátu částí, jako je intersepta.
Hemocelek: Hlavním tělním prostorem u členovců není coelom, ale krví naplněný hemocelek. Během embryonálního vývoje většiny členovců se v mezodermu alespoň některých somitů otevírají vestigiální coelomové dutiny. Ty jsou brzy obliterovány nebo jsou spojeny s prostorem mezi vyvíjejícími se mezodermálními a ektodermálními strukturami a žloutkem. Tento prostor se stává hemokolem, a není tedy vystlán mezodermálním peritoneem. U korýšů zůstávají jedinými coelomovými oddíly koncové váčky vylučovacích orgánů a prostor kolem gonád.
Svalová soustava: Podstatnou část těla většiny korýšů tvoří příčně pruhované svaly. Svaly jsou obvykle uspořádány do antagonistických skupin: flexory, které přitahují část těla k tělu, a extenzory, které ji prodlužují směrem ven. Břicho raků má silné ohýbače (obrázek 19-5), které se používají, když živočich plave dozadu – což je jeho nejlepší způsob úniku. Silné svaly na obou stranách břicha ovládají čelisti.
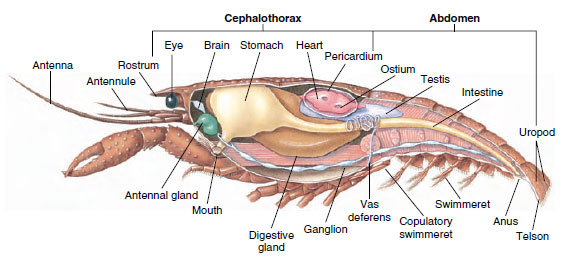
Obrázek 19-5
Vnitřní stavba samce raka.
Dýchací soustava: Výměna dýchacích plynů u menších korýšů probíhá na tenčích částech kutikuly (například v nohách) nebo na celém těle a specializované struktury mohou chybět. Větší korýši mají žábry, což jsou jemné, peříčkovité výběžky s velmi tenkou kutikulou. U desetinožců boky karapaxu uzavírají žaberní dutinu, která je vpředu a břichem otevřená (obr. 19-6). Žábry mohou vystupovat z pohrudniční stěny do žaberní dutiny, ze spojení hrudních končetin s tělem nebo z hrudních kostí. Poslední dva typy jsou typické pro raky. „Bailer“, část druhé horní čelisti, nasává vodu přes žaberní vlákna, do žaberní dutiny u základů nohou a ven ze žaberní dutiny na přední straně.
Oběhová soustava: Korýši a další členovci mají „otevřený“ nebo lakunární typ oběhové soustavy. To znamená, že nemají žíly a neoddělují krev od intersticiální tekutiny, jako je tomu u živočichů s uzavřenou soustavou. Hemolymfa (krev) opouští srdce prostřednictvím tepen, cirkuluje přes hemocelek a vrací se do žilních sinusů nebo prostorů namísto žil, než znovu vstoupí do srdce. Annelidé mají uzavřený systém, stejně jako obratlovci.
Hřbetní srdce je hlavním hnacím orgánem. Je to jednokomorový vak z příčně pruhované svaloviny. Hemolymfa vstupuje do srdce z okolní perikardiální dutiny párovými ostiemi s chlopněmi, které zabraňují zpětnému toku do dutiny (obr. 19-6). Ze srdce se hemolymfa dostává do jedné nebo více tepen. Chlopně v tepnách zabraňují zpětnému toku hemolymfy. Malé tepny se vyprazdňují do tkáňových dutin, které zase často ústí do velké hrudní dutiny (obrázek 19-6).
Odtud vedou aferentní kanály dutiny hemolymfu do žaber, pokud jsou přítomny, k výměně kyslíku a oxidu uhličitého. Hemolymfa se pak eferentními kanály vrací do perikardiální dutiny (obrázek 19-6).
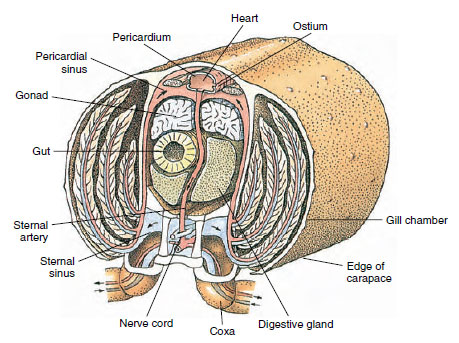
Obrázek 19-6
Diagramatický řez oblastí srdce raka ukazující směr toku krve v tomto „otevřeném“
krvním systému. Srdce pumpuje krev do tělesných tkání prostřednictvím tepen, které se vyprazdňují do tkáňových dutin. Vracející se
krev vstupuje do hrudní dutiny, pak prochází žábrami pro výměnu plynů a nakonec se vrací zpět do osrdečníkové dutiny eferentními
kanálky. Všimněte si absence žil.
Hemolymfa u členovců je převážně bezbarvá. Obsahuje ameboidní buňky nejméně dvou typů. V roztoku se může nacházet hemocanin, respirační pigment obsahující měď, nebo hemoglobin, pigment obsahující železo. Hemolymfa má vlastnost srážlivosti, která zabraňuje její ztrátě při drobných poraněních. Některé ameboidní buňky uvolňují koagulant podobný trombinu, který srážení sráží.
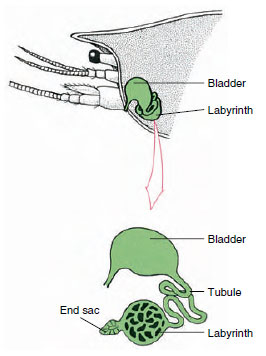
Obrázek 19-7
Schéma tykadlové žlázy (zelené žlázy)
raků. (V přirozené poloze je orgán značně
zalomený.) Některým korýšům chybí labyrint,
a vylučovací trubice (nefridiový kanál) je
velmi svinutá trubice.
Vylučovací soustava: Vylučovací orgány dospělých korýšů jsou dvojice trubicovitých struktur umístěných ve ventrální části hlavy před jícnem (obr. 19-5). Nazývají se tykadlové nebo čelistní žlázy podle toho, zda se otevírají na bázi tykadel nebo druhé čelisti. Několik dospělých korýšů má obě. Vylučovacími orgány desetinožců jsou tykadlové žlázy, v této skupině nazývané také zelené žlázy. Korýši nemají malpigické trubice, vylučovací orgány pavouků a hmyzu.
Koncový váček tykadlové žlázy, který pochází z embryonálního coelomového oddílu, se skládá z malého měchýřku (saccule) a houbovité hmoty zvané labyrint. Labyrint je spojen vylučovací trubicí s hřbetním měchýřkem, který se otevírá ven pórem na ventrálním povrchu bazálního článku tykadla (obrázek 19-7). Hydrostatický tlak v hemokoelu zajišťuje filtraci tekutiny do koncového vaku. Při průchodu filtrátu vylučovacími kanálky a močovým měchýřem dochází k jeho úpravě resorpcí solí, aminokyselin, glukózy a části vody a nakonec je vylučován jako moč.
Vylučování dusíkatých odpadních látek (většinou amoniaku) probíhá difuzí přes tenké plochy kutikuly, zejména přes žábry, a tzv. vylučovací orgány fungují hlavně pro regulaci iontového a osmotického složení tělních tekutin. Sladkovodní korýši, jako jsou raci, jsou neustále ohroženi nadměrným ředěním krve vodou, která difunduje přes žábry a jiné vodopropustné plochy. Zelené žlázy, které tvoří zředěnou moč s nízkým obsahem soli, fungují jako účinné „protipovodňové“ zařízení. Část Na+ a Cl- se ztrácí močí, ale tato ztráta je kompenzována aktivním vstřebáváním rozpuštěných solí žábrami. U mořských korýšů, jako jsou humři a krabi, ledviny upravují složení soli v hemolymfě selektivní úpravou obsahu soli v tubulární moči. U těchto forem zůstává moč izosmotická vůči krvi.
Nervová a smyslová soustava: Nervová soustava korýšů a annelidů má mnoho společného, i když u korýšů dochází k většímu splynutí ganglií (obr. 19-5). V mozku se nachází dvojice supraezofageálních ganglií, které zásobují nervy oči a dva páry tykadel. K němu se připojují spojky k subesofageálnímu gangliu, což je spojení nejméně pěti párů ganglií, které zásobují nervy ústa, přídatné orgány, jícen a tykadlové žlázy. Dvojitý ventrální nervový provazec má pár ganglií pro každý somit a nervy obsluhující přídatné orgány, svaly a další části.
Kromě tohoto centrálního systému může existovat sympatický nervový systém spojený s trávicím traktem.
Rakoještěři mají lépe vyvinuté smyslové orgány než annelidé. Největšími smyslovými orgány raků jsou oči a statocysty. Po těle jsou široce rozmístěny hmatové chloupky, jemné výběžky kutikuly, které jsou zvláště hojné na chelae, ústních částech a telsonu. Chemické smysly, chuť a čich, se nacházejí v chloupcích na tykadlech, ústních částech a dalších místech.
Na bazálním článku každého prvního tykadla raků se nachází vakovitá statocysta, která se otevírá na povrch hřbetním pórem. Statocysta obsahuje hřeben, na němž jsou smyslové chloupky vytvořené z chitinové výstelky a zrnka písku, která slouží jako statolity. Kdykoli živočich změní svou polohu, odpovídající změny polohy zrnek na smyslových chloupcích se přenášejí jako podněty do mozku a živočich se podle nich může přizpůsobit. Při každém stárnutí (ekdyze) kutikuly dochází ke ztrátě kutikulární výstelky statocysty a s ní i zrnek písku. Po ekdyse jsou hřbetním pórem nabrána nová zrna.
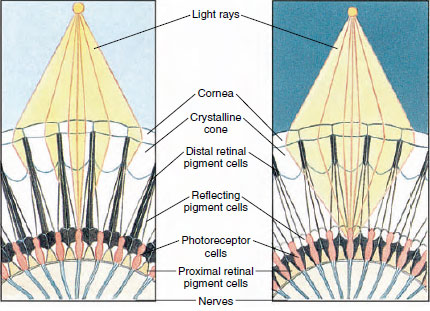
Obrázek 19-8
Část složeného oka členovce ukazující migraci pigmentu
v ommatidii pro denní a noční vidění. Na
každém obrázku je znázorněno pět ommatidií. Ve dne je každé ommatidium obklopeno tmavým
pigmentovým límcem, takže každé ommatidium je stimulováno pouze světelnými paprsky
, které vstupují do jeho vlastní rohovky (mozaikové vidění); v noci tvoří pigment
neúplné límce a světelné paprsky se mohou šířit do sousedních ommatidií
(souvislý nebo superpoziční obraz).
Oči u mnoha korýšů jsou složené, skládají se z mnoha fotoreceptorových jednotek zvaných ommatidia (obr. 19-8). Zaoblený povrch každého oka pokrývá průhledná plocha kutikuly, rohovka, která je rozdělena na mnoho malých čtverců nebo šestiúhelníků známých jako fasety. Tyto fasety jsou vnějšími konci ommatidií. Každé ommatidium se chová jako malé oko a obsahuje několik druhů buněk uspořádaných sloupcovitě (obrázek 19-8). Mezi sousedními ommatidii se nacházejí černé pigmentové buňky.
Pohyb pigmentu ve složeném oku členovců umožňuje přizpůsobit se různému množství světla. V každém ommatidiu jsou tři sady pigmentových buněk: distální retinální, proximální retinální a reflexní; ty jsou uspořádány tak, že mohou kolem každého ommatidia vytvořit více či méně úplný límec nebo rukáv. Při silné světelné nebo denní adaptaci se distální retinální pigment posouvá dovnitř a setkává se s proximálním retinálním pigmentem pohybujícím se směrem ven, takže se kolem ommatidia vytvoří úplný pigmentový rukáv (obrázek 19-8). Za tohoto stavu se k fotoreceptorovým buňkám (sítnici) dostanou pouze paprsky, které dopadají přímo na rohovku, protože každé ommatidium je chráněno před ostatními. Každé ommatidium tedy vidí pouze omezenou oblast zorného pole (mozaikový nebo apoziční obraz). Při slabém světle se distální a proximální pigmenty oddělují, takže světelné paprsky mají s pomocí reflexních pigmentových buněk možnost rozšířit se na sousední ommatidia a vytvořit souvislý nebo superpoziční obraz. Tento druhý typ vidění je méně přesný, ale maximálně využívá omezené množství přijímaného světla.
Reprodukce, životní cykly a endokrinní funkce
Většina korýšů má oddělená pohlaví a u různých skupin existují různé specializace na kopulaci. Střevlíci jsou jednodomí, ale obecně praktikují křížové oplození. U některých ostnokožců jsou samci vzácní a rozmnožování je obvykle partenogenetické. Většina korýšů líhne vajíčka nějakým způsobem: větvonožci a sudokopytníci mají speciální líhňařské komůrky, kopinatci mají líhňařské váčky připevněné na bocích břicha (viz obr. 19-19) a mnoho malakostrakanů nosí vajíčka a mláďata připevněná k břišním přívěskům.
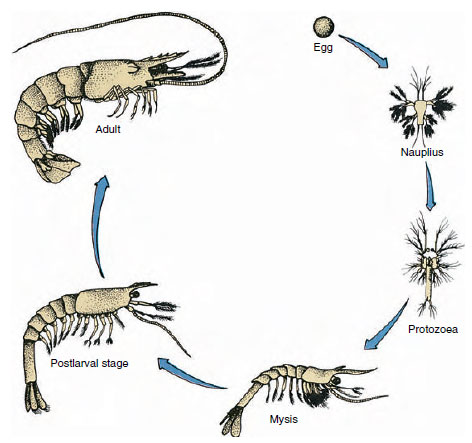
Obrázek 19-9
Životní cyklus garnáta Penaeus. Penaeidy se třou v hloubkách 40 až 90 m. Mladé larvální formy jsou
planktonní a přesouvají se na pobřeží do vody s nižší salinitou, kde se vyvíjejí jako juvenilní jedinci. Starší krevety se vracejí do hlubších vod
na pobřeží.
Rakovití mají přímý vývoj: neexistuje žádná larvální forma. Z vajíčka se vylíhne drobný juvenil, který má stejnou podobu jako dospělý jedinec a kompletní sadu přídatných orgánů a somitů. U většiny korýšů je však vývoj nepřímý a z vajíčka se vylíhne larva, která je svou stavbou a vzhledem zcela odlišná od dospělce. Přeměna larvy v dospělce je metamorfóza. Primitivní a nejrozšířenější larvou u korýšů je nauplius (obr. 19-9 a 19-23). Nauplie nesou pouze tři páry přídatných orgánů: uniramózní první tykadla, biramózní tykadla a biramózní čelisti. Všechny v tomto stádiu fungují jako plovací končetiny.
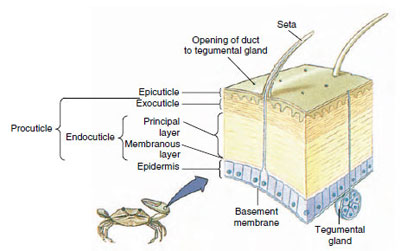
Obrázek 19-10
Struktura kutikuly korýšů.
Další vývoj může zahrnovat postupnou změnu k dospělé tělesné formě a přídatné orgány a somity se přidávají prostřednictvím řady svleček, nebo může převzetí dospělé formy zahrnovat náhlejší změny. Například metamorfóza mihulí probíhá od volně plovoucího nauplia k larvě s mlžím karapaxem zvaným cyprid a nakonec k přisedlému dospělci s vápenatými destičkami.
Ekdýza: Ekdýza (ek´duh-sis) (gr. ekdyein, svléknout) neboli pelichání je nezbytné pro zvětšení těla, protože exoskelet je neživý a s růstem živočicha neroste. Velká část fungování korýšů, včetně rozmnožování, chování a mnoha metabolických procesů, je přímo ovlivněna fyziologií cyklu pelichání.
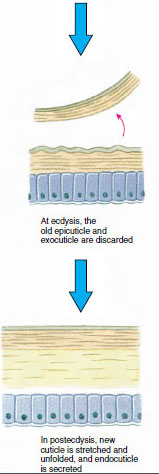
Kutikula, která je vylučována podkladovou epidermis, má několik vrstev (obr. 19-10). Nejzevnější je epikutikula, velmi tenká vrstva bílkovin napuštěných lipidy. Většinu kutikuly tvoří několik vrstev prokutikuly: (1) exokutikula, která je těsně pod epikutikulou a obsahuje bílkoviny, vápenaté soli a chitin; (2) endokutikula, která se sama skládá z (3) hlavní vrstvy, která obsahuje více chitinu a méně bílkovin a je silně kalcifikovaná, a (4) nevápenaté membránové vrstvy, poměrně tenké vrstvy chitinu a bílkovin.
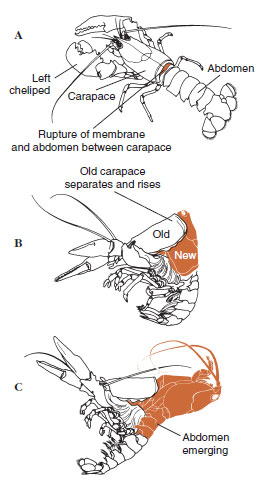
Nějakou dobu před vlastní ekdyzou se epidermální buňky značně zvětší. Oddělí se od membránové vrstvy, vylučují novou epikutikulu a začnou vylučovat novou exokutikulu (obr. 19-11). Do oblasti nad novou epikutikulou se uvolňují enzymy. Tyto enzymy začnou rozpouštět starou endokutikulu a rozpustné produkty se resorbují a ukládají v těle korýše. Některé soli vápníku se ukládají jako gastrolity (minerální nánosy) ve stěnách žaludku. Nakonec zůstane ze staré kutikuly pouze exokutikula a epikutikula, které jsou podloženy novou epikutikulou a novou exokutikulou. Zvíře polyká vodu, kterou vstřebává střevem, a výrazně se zvětšuje objem jeho krve. Vnitřní tlak způsobí, že se kutikula rozštěpí a živočich se vytáhne ze starého exoskeletu (obrázek 19-12). Následuje natažení dosud měkké nové kutikuly, usazení nové endokutikula, opětovné usazení zachráněných anorganických solí a dalších složek a zpevnění nové kutikuly. Během období pelichání je živočich bezbranný a zůstává skrytý.
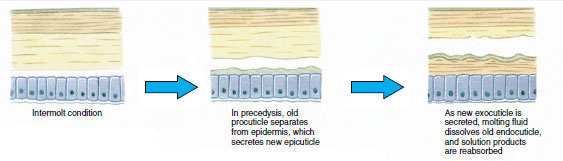 |
|
 |
 |
| Obrázek 19-12 Sekvence pelichání u humra Homarus americanus. A, Membrána mezi karapaxem a břichem praskne a karapax se začne pomalu zvedat. Tento krok může trvat až 2 hodiny. B a C, hlava, hrudník a nakonec břicho se stáhnou. Tento proces obvykle netrvá déle než 15 minut. Bezprostředně po ekdyse jsou čelisti vysušené a tělo je velmi měkké. Humr pokračuje v rychlém vstřebávání vody, takže během 12 hodin se tělo zvětší asi o 20 % na délku a 50 % na hmotnost. Tkáňová voda bude v následujících týdnech nahrazena bílkovinami. |
Obrázek 19-11 Vylučování a resorpce kůže při ekdyse. |
Když je korýš mladý, musí k ekdyse docházet často, aby mohl růst, a cyklus línání je relativně krátký. Jak se živočich blíží k dospělosti, periody intermolt se postupně prodlužují a u některých druhů se pelichání zcela zastaví. Během období intermolt dochází k nárůstu tkáňové hmoty, protože živá tkáň nahrazuje vodu.
Hormonální řízení cyklu ekdyzy
Ačkoli je ekdyza řízena hormonálně, cyklus je často iniciován podnětem z prostředí, který vnímá centrální nervová soustava. Takovými podněty mohou být teplota, délka dne a vlhkost (u suchozemských krabů). Signál z centrální nervové soustavy snižuje produkci hormonu inhibujícího tání v X-orgánu. X-organ je skupina neurosekrečních buněk v medulla terminalis mozku. U raků a dalších desetinožců se medulla terminalis nachází v oční stopce. Hormon je přenášen axony X-orgánu do sinusové žlázy (která sama pravděpodobně nemá žlázovou funkci), rovněž v oční stopce, kde je uvolňován do hemolymfy.
Pokles hladiny hormonu vyvolávajícího línání podporuje uvolňování hormonu vyvolávajícího línání z Y-orgánu. Yorgany jsou pod epidermis v blízkosti přitahovacích svalů čelistí a jsou homol-ogusní s prothorakálními žlázami hmyzu, které produkují hormon ekdyson. Působení hormonu pelichání spočívá v zahájení procesů vedoucích k ekdyse (proekdyse). Jakmile je cyklus zahájen, probíhá automaticky bez dalšího působení hormonů z orgánů X nebo Y.
Další endokrinní funkce: Nejenže odstranění očních stopek urychluje línání, ale před více než 100 lety bylo také zjištěno, že korýši, kterým byly odstraněny oční stopky, již nemohou přizpůsobit zbarvení těla podmínkám pozadí. Před více než 50 lety bylo zjištěno, že tato vada není způsobena ztrátou zraku, ale ztrátou hormonů v očních stopkách. Zbarvení těla korýšů je z velké části výsledkem působení pigmentů ve speciálních rozvětvených buňkách (chromatoforech) v epidermis.
Koncentrace pigmentových granulí ve středu buněk způsobuje zesvětlující efekt a rozptýlení pigmentu po celých buňkách způsobuje ztmavující efekt. Chování pigmentu je řízeno hormony z neurosekrečních buněk v oční stopce, stejně jako migrace sítnicového pigmentu pro adaptaci na světlo a tmu v očích (obr. 19-8).
Uvolňování neurosekrečního materiálu z osrdečníkových orgánů ve stěně osrdečníku způsobuje zvýšení frekvence a amplitudy srdečního tepu.
Androgenní žlázy, poprvé nalezené u obojživelníka (Orchestia, běžný plážový skokan), se vyskytují u samců malakostrakanů. Na rozdíl od většiny ostatních endokrinních orgánů u korýšů se nejedná o neurosekreční orgány. Jejich sekrece stimuluje projevy samčích pohlavních znaků. Mladí malakostrakani mají rudimentární androgenní žlázy, ale u samic se tyto žlázy nevyvinou. Pokud jsou samici uměle implantovány, její vaječníky se přemění na varlata a začnou produkovat spermie a její přívěsky začnou při dalším línání nabývat samčích znaků. U izopodů se androgenní žlázy nacházejí ve varlatech, u všech ostatních malakostrakanů jsou mezi svaly koxopodů posledních hrudních končetin a částečně připojeny poblíž konců vasa deferentia. Samice sice nemají orgány podobné androgenním žlázám, ale jejich vaječníky produkují jeden nebo dva hormony, které ovlivňují sekundární pohlavní znaky.
U korýšů mohou být přítomny hormony, které ovlivňují další tělesné pochody, a existují důkazy, že neurosekreční látka produkovaná v oční stopce reguluje hladinu cukru v krvi.
Životní návyky
Životní návyky a adaptace na krmení se u jednotlivých korýšů značně liší. Mnoho forem může přecházet z jednoho typu krmení na jiný v závislosti na prostředí a dostupnosti potravy, ale všechny používají stejný základní soubor ústních ústrojí. Čelisti a horní čelisti slouží k polykání potravy; horní čelisti potravu drží a drtí. U dravců slouží k zachycování potravy kráčivé končetiny, zejména chelipedy.
Mnoho korýšů, velkých i malých, je dravých a někteří mají zajímavé adaptace k usmrcování kořisti. Jedna z krevetovitých forem, Lygiosquilla, má na jedné z kráčivých nohou specializovaný prst, který lze vtáhnout do žlábku a náhle uvolnit, aby probodl procházející kořist. Pistolové krevety (Alpheus spp.) mají obrovsky zvětšenou chelu, která se dá natáhnout jako kladívko pistole a udeřit silou, která kořist omráčí.
Potrava závěsníků sahá od planktonu a detritu až po bakterie. Dravci konzumují larvy, červy, korýše, plže a ryby. Mrchožrouti se živí odumřelou živočišnou a rostlinnou hmotou. Závěsní krmiči, jako jsou krevetky, vodní blechy a mlži, používají své nohy s hustými třásněmi, které vytvářejí vodní proudy, jimiž jsou částečky potravy unášeny. Bahenní krevety (Upogebia spp.) používají dlouhé setae na prvních dvou párech hrudních přívěsků k odčerpávání potravy z vody, která cirkuluje v jejich noře pohyby jejich plováků.
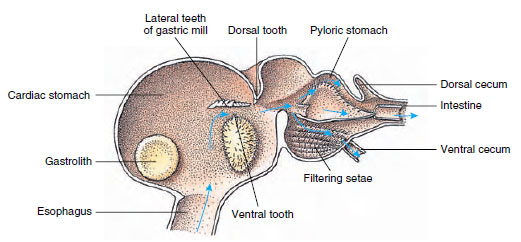
Rakovití mají dvoudílný žaludek (obr. 19-13). První část obsahuje žaludeční mlýnek, v němž může být potrava, již roztrhaná čelistmi, dále rozmělněna třemi vápenatými zuby na částice dostatečně jemné, aby prošly setózovým filtrem v druhé části; částice potravy pak přecházejí do střeva k chemickému trávení.
Obrázek 19-13
Žaludek malakostrakana ukazuje žaludeční „mlýnek“ a směry pohybu potravy. Mlýnek má chitinové hřebeny neboli
zuby pro žvýkání a setae pro cedění potravy před přechodem do pylorického žaludku
.
Napsat komentář