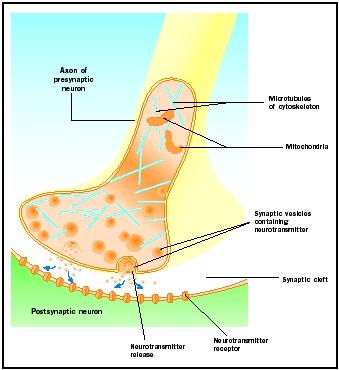
Frisättning av neurotransmittor
Aktionspotentialer som når synaptiska knoppar utlöser frisättning av neurotransmittor i den synaptiska klyftan. Den molekylära mekanismen är inte helt klarlagd. En ”synaptisk fördröjning” på en till två millisekunder inträffar mellan ankomsten av aktionspotentialen och frisättningen av neurotransmittorn. Handlingspotentialen öppnar kalciumkanaler i den synaptiska knölens membran, vilket orsakar en inåtriktad rörelse av kalciumjoner . Kalciumjonerna utlöser frisättning av neurotransmittor från synaptiska vesiklar i den synaptiska klyftan. De synaptiska vesiklarna smälter samman med det presynaptiska membranet under denna exocytosprocess. De gamla vesiklarnas membran blir en del av det presynaptiska membranet och nya vesiklar klämmer av sig från ett angränsande membranområde. Dessa nya vesiklar fylls sedan på med nysyntetiserade eller ”återvunna” neurotransmittorer.
De frigjorda neurotransmittorerna diffunderar över den smala synaptiska klyftan. Vid det postsynaptiska membranet binder neurotransmittormolekyler till membranbundna receptormolekyler med igenkänningsställen som är specifika för den neurotransmittorn. Neurotransmittorns bindning till receptorn utlöser ett postsynaptiskt svar som är specifikt för den receptorn. Dessa reaktioner kan vara antingen excitatoriska eller hämmande, beroende på receptorns egenskaper. Om receptorstimulering leder till att det postsynaptiska membranet blir mer elektriskt positivt (depolariserat) är det en excitatorisk postsynaptisk potential (EPSP). Om det blir mer negativt (hyperpolariserat) är det en hämmande postsynaptisk potential (IPSP). Excitering och hämning beror på receptorns egenskaper och inte på neurotransmittorn. Receptorer som är kopplade till natrium- eller kalciumkanaler är excitatoriska och ger upphov till en depolarisering av det postsynaptiska membranet, medan receptorer som är kopplade till klorid- eller kaliumkanaler är hämmande och ger upphov till en hyperpolarisering av det postsynaptiska membranet. Sådana receptorer som är kopplade till jonkanaler kallas jonotropa receptorer.
Andra receptorer är kopplade till ”second-messenger”-system som initierar en serie biokemiska reaktioner i den postsynaptiska cellen. Dessa är metabotropa receptorer . Metabotropa receptorer kan producera många olika
postsynaptiska händelser. Dessa sträcker sig från direkt aktivering av intilliggande jonkanaler, till förändring av receptorkänsligheten, till transkription av specifika budbärarribonukleinsyror (RNA) eller till och med aktivering av specifika gener . Kemiska synapser är en del av ett mycket anpassningsbart och flexibelt kommunikationssystem. Det är inte statiska anatomiska strukturer med fasta egenskaper utan dynamiska strukturer som kan ändra sina molekylära egenskaper under förändrade omständigheter.
Det finns bokstavligen hundratals neurotransmittorer. Vissa är ganska enkla föreningar som acetylkolin, serotonin, katekolaminerna (dopamin, noradrenalin och adrenalin) och ett antal aminosyror . Många är mer komplexa och tillhör det stora antalet neuropeptidtransmittorer. När de väl har släppts ut i den synaptiska klyftan förblir neurotransmittorerna aktiva tills de antingen förändras kemiskt eller tas tillbaka till den synaptiska knoppen av särskilda transportsystem och återvinns. Vid kolinerga synapser finns acetylkolinesteras i den synaptiska klyftan. Detta enzym klyver neurotransmittorn till acetat och kolin, som båda inte är aktiva. Serotonin och adrenalin däremot tas upp i den presynaptiska terminalen och återvinns.
Lämna ett svar