Subphylum Crustacea
General Nature of a Crustace
Crustaceaner skiljer sig från andra leddjur på många olika sätt, men det enda som verkligen skiljer dem från andra leddjur är att de är de enda leddjuren som har två par antenner. Förutom två par antenner och ett par mandibler har kräftdjur två par maxiller på huvudet, följt av ett par bihang på varje kroppssegment eller somit. Hos vissa kräftdjur är inte alla somiter försedda med bihang. Alla bihang, utom kanske de första antennerna, är primitivt biramösa (två huvudgrenar), och åtminstone vissa bihang hos dagens vuxna djur uppvisar detta tillstånd. Organ som är specialiserade på andning, om de finns, fungerar som gälar.
De flesta kräftdjur har mellan 16 och 20 somiter, men vissa former har 60 somiter eller mer. Ett större antal somiter är ett primitivt kännetecken. Det mer härledda tillståndet är att ha färre segment och ökad tagmatisering. Större tagmata är huvud, thorax och buk, men dessa är inte homologa i hela klassen (eller till och med inom vissa underklasser) på grund av varierande grad av sammansmältning av somiter, till exempel som i cephalothorax.
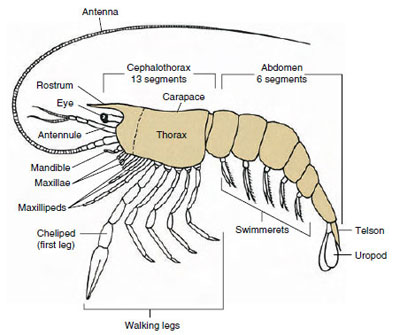
Figur 19-1
Arketypisk plan över Malacostraca. De två maxillerna och de tre
maxillipederna har separerats schematiskt för att illustrera
den
generella planen.
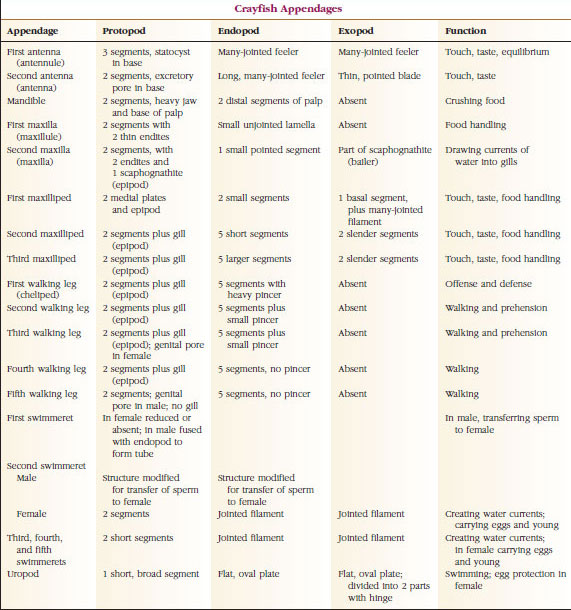
Den överlägset största gruppen kräftdjur är klassen Malacostraca, som innefattar hummer, krabbor, räkor, strandhoppare, sengångare och många andra. Dessa uppvisar ett förvånansvärt konstant arrangemang av kroppssegment och tagmata, vilket anses vara klassens förfäders plan (figur 19-1). Denna typiska kroppsplan har ett huvud med fem (sex embryonalt) sammansmälta somiter, ett thorax med åtta somiter och en buk med sex somiter (sju hos några få arter). I den främre änden finns ett icke-segmenterat rostrum och i den bakre änden finns ett icke-segmenterat telson, som tillsammans med den sista buksomiten och dess uropoder bildar en svansfläkt i många former.
I många kräftdjur kan huvudets dorsala kutikula sträcka sig bakåt och runt om sidorna av djuret för att täcka eller smälta samman med några eller alla bröst- och buksomiter. Denna täckning kallas för karapax. Hos vissa grupper bildar karapaxen musselliknande klaffar som täcker större delen av eller hela kroppen. Hos decapoder (inklusive hummer, räkor, kräftor och andra) täcker karapaxet hela cephalothorax men inte buken.
Form och funktion
På grund av deras storlek och lättillgänglighet har stora kräftdjur som kräftor studerats mer än andra grupper. De studeras också ofta i introducerande laboratoriekurser. Därför gäller många av kommentarerna som följer specifikt för kräftor och deras släktingar.
Uttre egenskaper
Kropparna hos kräftor är täckta av en utsöndrad kutikula som består av kitin, protein och kalkhaltigt material. De hårdare, tunga plattorna hos större kräftdjur har ett särskilt högt innehåll av kalkavlagringar. Det hårda skyddande höljet är mjukt och tunt vid lederna mellan somiterna, vilket möjliggör flexibla rörelser. Carapacen, om den finns, täcker en stor del av eller hela cephalothorax; hos decapoder som kräftor är alla huvud- och bröstsegment dorsalt omslutna av carapacen. Varje somit som inte är omsluten av pannan är täckt av en dorsal kutikulär platta, eller tergum (figur 19-2A), och en ventral tvärgående bar, sternum, ligger mellan segmentens bilagor (figur 19-2B). Buken avslutas med ett telson, som inte betraktas som en somit och som bär anus. (Telsonet kan vara homologt till annelidernas pygidium.)
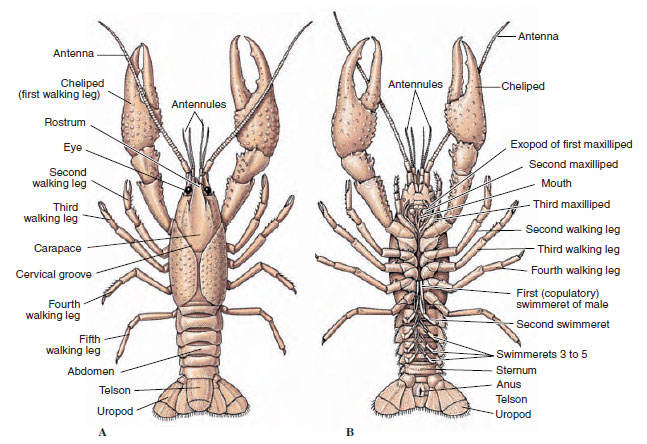
Figur 19-2
Kräftors yttre struktur. A, Dorsal vy. B, Ventralvy.
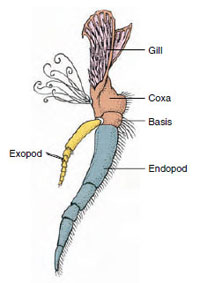
Figur 19-3
Delar av ett biramöst kräftdjur
tillbehör (tredje maxillipeden hos en
kräfta).
Gonoporernas placering varierar beroende på kön och grupp av kräftdjur. De kan sitta på eller vid basen av ett par bihang, vid kroppens ändpunkt eller på somiter utan ben. Hos kräftor ligger vasa deferentias öppningar på mediansidan vid basen av det femte paret gångben, och äggledarnas öppningar vid basen av det tredje paret. Hos honorna är öppningen till sädesbehållaren vanligen belägen i midventrallinjen mellan det fjärde och femte paret gångben.
Apparater: Medlemmar av klasserna Malacostraca (inklusive kräftor) och Remipedia har vanligtvis ett par ledade bihang på varje somit (figur 19-3), även om buksomiterna i de andra klasserna inte bär bihang. En avsevärd specialisering är uppenbar i bihang hos avledda kräftdjur som kräftor. Alla är dock variationer av den grundläggande, tvåformiga planen, som illustreras av ett kräftans bihang, t.ex. en maxilliped (en bröstkorgsben som modifierats för att bli ett huvudtillbehör) (figurerna 19-3 och 19-4). Den basala delen, eller protopoden, har en lateral exopod och en medial endopod. Protopoden består av en eller två leder (bas och coxa), medan exopoden och endopoden har en eller flera leder vardera. Vissa bihang, som t.ex. gåben hos flodkräftor, har blivit sekundärt uniförbundna. Mediala eller laterala processer förekommer ibland på kräftdjurens lemmar, som kallas endites respektive exites, och en exite på protopoden kallas epipod. Epipoderna är ofta modifierade som gälar. Tabell 19-1 visar hur de olika bihangenserna har modifierats från den biramösa planen för att passa specifika funktioner.
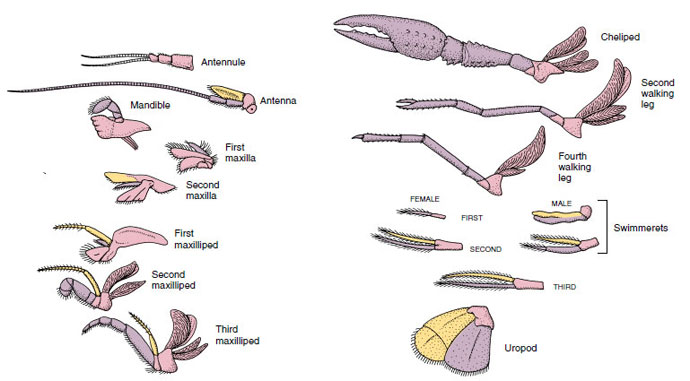
Figur 19-4
Anhangsdelar hos en flodkräfta som visar hur de har blivit modifierade från den grundläggande biramösa planen, så som de återfinns hos en
simmare. Protopod, rosa; endopod, lila; exopod, gul.
Strukturer som har en liknande grundplan och som härstammar från en gemensam form sägs vara homologa, oavsett om de har samma funktion eller inte. Eftersom specialiserade gångben, munhålor, chelipeder och simmande ben alla har utvecklats från en gemensam biramös typ, men har modifierats för att utföra olika funktioner, är de alla homologa till varandra, ett tillstånd som kallas seriell homologi. Primitivt var lemmar alla mycket lika, men under evolutionen av strukturella modifieringar har vissa grenar reducerats, vissa försvunnit, vissa ändrats kraftigt och vissa nya delar har lagts till. Kräftor och deras allierade har den mest genomarbetade seriella homologin i djurriket och har 17 olika men seriellt homologa typer av bihang (tabell 19-1).
TABELL 19.1
Interna kännetecken
Muskel- och nervsystemet samt segmenteringen i thorax och buk visar tydligt på metamerism som ärvts från annelidliknande förfäder, men det finns tydliga modifieringar i andra system. De flesta förändringar innebär att delar koncentreras till ett visst område eller att delar minskas eller helt försvinner, t.ex. intersepta.
Hemocoel: Det största kroppsutrymmet hos leddjuren är inte coelom utan en blodfylld hemocoel. Under embryonalutvecklingen hos de flesta leddjur öppnas rudimentära coelomiska hålrum i mesodermen hos åtminstone vissa somiter. Dessa är snart utplånade eller blir sammanhängande med utrymmet mellan de mesodermala och ektodermala strukturerna under utveckling och äggulan. Detta utrymme blir hemocoel och är alltså inte fodrat av ett mesodermiskt peritoneum. Hos kräftdjur är de enda coelomiska avdelningarna som återstår de utsöndringsorganens ändsäckar och utrymmet runt könskörtlarna.
Muskulärt system: De flesta kräftdjur har en betydande del av kroppen uppbyggd av streckade muskler. Musklerna är vanligtvis ordnade i antagonistiska grupper: flexorer, som drar en del mot kroppen, och extensorer, som sträcker den utåt. Kräftans buk har kraftiga böjare (figur 19-5), som används när djuret simmar bakåt – dess bästa sätt att fly. Starka muskler på vardera sidan av magen styr mandiblerna.
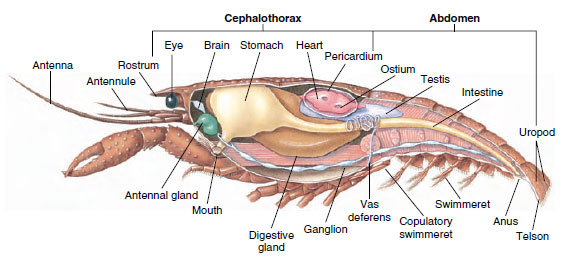
Figur 19-5
Inre struktur hos en manlig kräfta.
Respiratoriskt system: Andningsgasutbytet hos mindre kräftdjur sker över tunnare områden av kutikulan (till exempel i benen) eller över hela kroppen, och specialiserade strukturer kan saknas. Större kräftdjur har gälar, som är känsliga, fjäderliknande utsprång med mycket tunn kutikula. Hos decapoderna omsluter sidorna av karapaxet gälhålan, som är öppen framifrån och ventralt (figur 19-6). Gälarna kan skjuta ut från pleuraväggen in i gälhålan, från bröstbenens artikulation med kroppen eller från bröstbenen. De två sistnämnda typerna är typiska för kräftor. ”Bailer”, en del av den andra överkäken, drar vatten över gältrådarna, in i gälhålan vid benens baser och ut ur gälhålan vid den främre.
Cirkulationssystemet: Kräftdjur och andra leddjur har ett ”öppet” eller lacunärt cirkulationssystem. Detta innebär att det inte finns några vener och ingen separation av blodet från interstitiell vätska, vilket är fallet hos djur med slutna system. Hemolymfen (blodet) lämnar hjärtat via artärer, cirkulerar genom hemocoelen och återvänder till venösa bihålor, eller utrymmen, i stället för vener innan den återvänder till hjärtat. Annelider har ett slutet system, liksom ryggradsdjur.
Ett rygghjärta är det huvudsakliga drivorganet. Det är en enkammarsäck av strimmiga muskler. Hemolymfen kommer in i hjärtat från den omgivande perikardiella sinus genom parade ostior, med ventiler som förhindrar återflöde in i sinus (figur 19-6). Från hjärtat kommer hemolymphen in i en eller flera artärer. Ventiler i artärerna förhindrar ett återflöde av hemolymphe. Små artärer mynnar ut i vävnadssinus, som i sin tur ofta mynnar ut i en stor sinus sternalis (figur 19-6).
Därifrån transporterar afferenta sinuskanaler hemolymphe till gälarna, om sådana finns, för utbyte av syre och koldioxid. Hemolymfen återvänder sedan till sinus pericardial genom efferenta kanaler (figur 19-6).
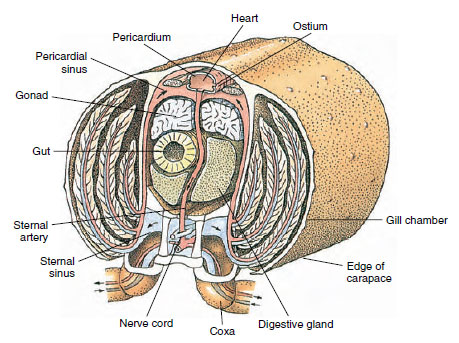
Figur 19-6
Diagrammatiskt tvärsnitt genom hjärtregionen hos en flodkräfta som visar riktningen på blodflödet i detta ”öppna”
blodsystem. Hjärtat pumpar blod till kroppens vävnader genom artärer som mynnar ut i vävnadssinus. Det återkommande
blodet går in i sinus sternalis, går sedan genom gälarna för gasutbyte och slutligen tillbaka till sinus pericardialis genom efferenta
kanaler. Notera avsaknaden av vener.
Hemolymphe hos leddjur är i stort sett färglös. Den innehåller ameboida celler av minst två typer. Hemocyanin, ett kopparhaltigt andningspigment, eller hemoglobin, ett järnhaltigt pigment, kan finnas i lösning. Hemolymfen har egenskapen att koagulera, vilket förhindrar att den förloras vid mindre skador. Vissa ameboidceller avger ett trombinliknande koaguleringsmedel som fäller ut koaguleringen.
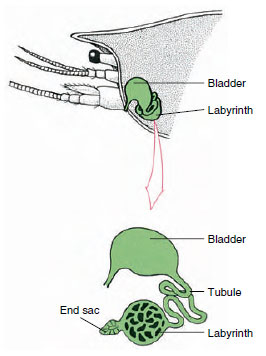
Figur 19-7
Skema av antennkörteln (grön körtel) hos
kräftor. (I naturligt läge är organet mycket
veckat.) Vissa kräftdjur saknar labyrint,
och utsöndringsröret (nephridialkanalen) är
ett mycket hoprullat rör.
Utsöndringssystemet: Utsöndringsorganen hos vuxna kräftdjur är ett par tubulära strukturer som är belägna i den ventrala delen av huvudet framför matstrupen (Figur 19-5). De kallas antennkörtlar eller maxillarkörtlar, beroende på om de öppnas vid basen av antennerna eller vid basen av de andra maxillarna. Några få vuxna kräftdjur har båda. Utsöndringsorganen hos decapoder är antennkörtlar, även kallade gröna körtlar i den här gruppen. Kräftdjur har inte malpighianska tubuli, spindlars och insekters utsöndringsorgan.
Antennalkörtelns ändsäck, som härstammar från ett embryonalt coelomiskt fack, består av en liten vesikel (saccule) och en svampig massa som kallas labyrint. Labyrinten ansluter genom en utsöndringstubuli till en dorsal blåsa, som öppnas utåt genom en por på den ventrala ytan av det basala antennsegmentet (figur 19-7). Det hydrostatiska trycket i hemocoelen ger kraft för filtrering av vätska in i ändsäcken. När filtratet passerar genom utsöndringstubuli och urinblåsan modifieras det genom resorption av salter, aminosyror, glukos och en del vatten och utsöndras slutligen som urin.
Utsöndring av kvävehaltigt avfall (främst ammoniak) sker genom diffusion genom tunna områden av kutikulan, särskilt gälarna, och de s.k. utsöndringsorganen fungerar huvudsakligen för att reglera den joniska och osmotiska sammansättningen av kroppsvätskor. Sötvattenskräftor, t.ex. kräftor, hotas ständigt av en överspädning av blodet med vatten, som diffunderar genom gälarna och andra vattengenomsläppliga ytor. De gröna körtlarna, som bildar en utspädd, saltfattig urin, fungerar som en effektiv anordning för att kontrollera översvämningar. En del Na+ och Cl- försvinner i urinen, men denna förlust kompenseras genom att gälarna aktivt absorberar löst salt. Hos marina kräftdjur, t.ex. hummer och krabba, fungerar njurarna för att justera saltsammansättningen i hemolymfen genom att selektivt modifiera saltinnehållet i den tubulära urinen. I dessa former förblir urinen isosmotisk i förhållande till blodet.
Nerv- och sinnessystem: Nervsystemen hos kräftdjur och anneldjur har mycket gemensamt, även om kräftdjurens nervsystem har fler fusioner av ganglier (figur 19-5). Hjärnan är ett par supraesophageala ganglier som förser nerver till ögonen och två par antenner. Den är sammanfogad genom bindemedel med det subesofageala gangliet, en sammanslagning av minst fem par ganglier som förser munnen, bihang, matstrupen och antennkörtlarna med nerver. Den dubbla ventrala nervsträngen har ett par ganglier för varje somit och nerver som försörjer bihang, muskler och andra delar.
Förutom detta centrala system kan det finnas ett sympatiskt nervsystem som är förknippat med matsmältningskanalen.
Kräftdjur har bättre utvecklade sinnesorgan än anneldjur. De största sinnesorganen hos kräftor är ögon och statocystor. Över hela kroppen finns känselhår, känsliga utskjutningar av kutikula som är särskilt rikligt förekommande på chelae, munhålor och telson. De kemiska sinnena smak och lukt finns i hår på antenner, mundelar och andra ställen.
En säckliknande statocysta, som öppnar sig mot ytan genom en dorsal por, finns på det basala segmentet av varje första antenn hos kräftor. Statocystan innehåller en kam som bär på känselhår som bildats av det kitinhaltiga fodret och sandkorn som fungerar som statoliter. När djuret ändrar sin position, skickas motsvarande förändringar i positionen för kornen på känselhåren som stimuli till hjärnan, och djuret kan anpassa sig i enlighet med detta. Vid varje skiftning (ecdysis) av kutikulan förloras statocystans kutikulära foder och med det sandkornen. Nya korn plockas upp genom dorsalporten efter ecdysis.
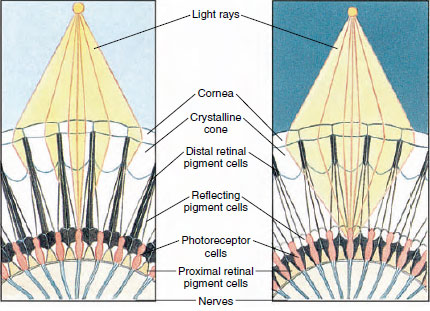
Figur 19-8
Del av sammansatt öga hos en ledfot som visar migrationen av pigment
i ommatidia för dag- och nattsyn. Fem ommatidier är representerade i
vardera diagrammet. På dagen är varje ommatidium omgivet av en mörk
pigmentkrage så att varje ommatidium stimuleras endast av ljusstrålar
som kommer in i den egna hornhinnan (mosaikseende); på natten bildar pigmentet
inkompletta kragar och ljusstrålar kan sprida sig till intilliggande ommatidia
(kontinuerlig, eller superposition, bild).
Ögonen hos många kräftdjur är sammansatta och består av många fotoreceptorenheter som kallas ommatidia (figur 19-8). Den rundade ytan på varje öga täcks av ett genomskinligt område av kutikula, hornhinnan, som är uppdelad i många små fyrkanter eller hexagoner som kallas facetter. Dessa facetter är de yttre ändarna av ommatidierna. Varje ommatidium beter sig som ett litet öga och innehåller flera sorters celler som är anordnade på ett kolonnformigt sätt (figur 19-8). Svarta pigmentceller finns mellan intilliggande ommatidia.
Pigmentens rörelse i ett leddjurens sammansatta öga gör att det kan anpassa sig till olika mängder ljus. Det finns tre uppsättningar pigmentceller i varje ommatidium: distala retinala, proximala retinala och reflekterande; dessa är så arrangerade att de kan bilda en mer eller mindre komplett krage eller hylsa runt varje ommatidium. Vid stark ljus- eller daganpassning rör sig det distala retinalpigmentet inåt och möter det proximala retinalpigmentet som rör sig utåt, så att det bildas en komplett pigmenthylsa runt ommatidium (figur 19-8). I detta tillstånd kommer endast strålar som träffar hornhinnan direkt att nå fotoreceptorcellerna (retinularcellerna), eftersom varje ommatidium är skyddat från andra. Således kommer varje ommatidium endast att se ett begränsat område av synfältet (en mosaik- eller appositionsbild). I svagt ljus separeras distala och proximala pigment så att ljusstrålarna, med hjälp av reflekterande pigmentceller, har en chans att sprida sig till intilliggande ommatidier och bilda en sammanhängande bild, eller superpositionsbild. Denna andra typ av syn är mindre exakt men utnyttjar maximalt den begränsade mängd ljus som tas emot.
Reproduktion, livscykler och endokrin funktion
De flesta kräftdjur har separata kön, och det finns olika specialiseringar för kopulation bland olika grupper. Snäckor är enäggiga men praktiserar i allmänhet korsbefruktning. Hos vissa ostrakoder är hanar sällsynta, och reproduktionen är vanligen parthenogenetisk. De flesta kräftdjur ruvar sina ägg på något sätt: grenlöss och havstulpaner har särskilda yngelkammare, copepoder har yngelkammare som är fästa på sidorna av buken (se figur 19-19), och många malakostrakier bär ägg och ungar fästa på sina buktillbehör.
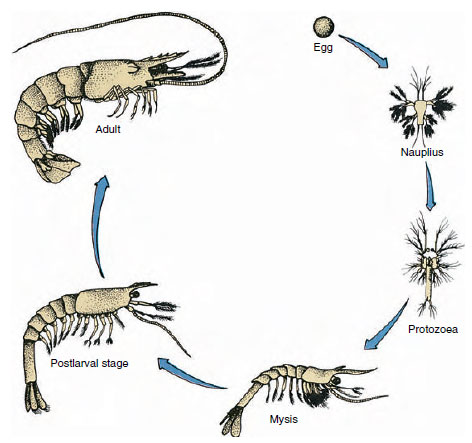
Figur 19-9
Livscykel hos en gulfkärrsräka Penaeus. Penaeiderna leker på 40-90 meters djup. De unga larvformerna är
planktoniska och rör sig intill kusten till vatten med lägre salthalt för att utvecklas som juvenila individer. Äldre räkor återvänder till djupare vatten
ut till land.
Kräftor har en direkt utveckling: det finns ingen larvform. En liten juvenil med samma form som den vuxna och en komplett uppsättning bihang och somiter kläcks ur ägget. Utvecklingen är dock indirekt hos de flesta kräftdjur, och en larv som är helt olik den vuxne till struktur och utseende kläcks ur ägget. Förändringen från larv till vuxen är metamorfos. Den primitiva och mest förekommande larven hos kräftdjuren är nauplius (figur 19-9 och 19-23). Nauplierna har endast tre par bilagor: uniramösa första antenner, biramösa antenner och biramösa mandibler. Samtliga fungerar som simföremål i detta skede.
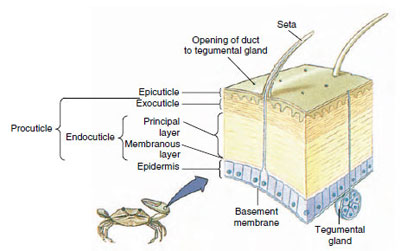
Figur 19-10
Kräftdjurens kutikulära struktur.
Den efterföljande utvecklingen kan innebära en gradvis förändring till den vuxna kroppsformen, och bihang och somiter läggs till genom en serie av skiftningar, eller så kan antagandet av den vuxna formen innebära mer abrupta förändringar. Metamorfosen hos en havstulpan går till exempel från en fritt simmande nauplius till en larv med ett tvåskaligt skal som kallas cyprid och slutligen till en fastsittande vuxen med kalkplattor.
Ecdysis: Ecdysis (ek´duh-sis) (Gr. ekdyein, att ta av sig), eller molting, är nödvändigt för att kroppen ska kunna öka i storlek eftersom exoskelettet inte lever och inte växer i takt med att djuret växer. Mycket av ett kräftdjurs funktionssätt, inklusive reproduktion, beteende och många metaboliska processer, påverkas direkt av den fysiologi som hör samman med ruggningscykeln.
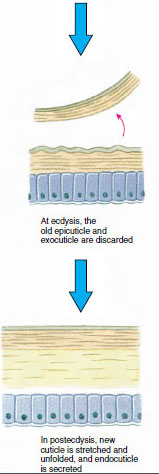
Hudskivan, som utsöndras av den underliggande epidermis, har flera lager (figur 19-10). Det yttersta är epicuticle, ett mycket tunt lager av lipidimpregnerat protein. Huvuddelen av kutikulan utgörs av flera lager procuticle: (1) exocuticle, som ligger precis under epicuticle och innehåller protein, kalciumsalter och kitin, (2) endocuticle, som i sin tur består av (3) ett huvudskikt, som innehåller mer kitin och mindre protein och är kraftigt förkalkat, och (4) ett okalkat membranskikt, ett relativt tunt skikt av kitin och protein.
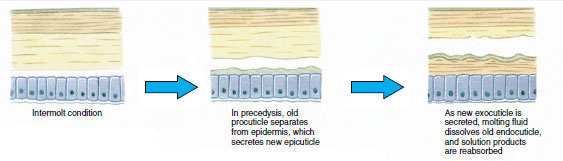
En bit före den egentliga ecdysen förstorar epidermalcellerna sig avsevärt. De separerar sig från det membranösa lagret, utsöndrar en ny epicuticle och börjar utsöndra en ny exocuticle (figur 19-11). Enzymer frigörs i området ovanför den nya epiktikeln. Dessa enzymer börjar lösa upp den gamla endokutikeln, och de lösliga produkterna resorberas och lagras i kräftdjurets kropp. Vissa kalciumsalter lagras som gastroliter (mineralansamlingar) i magsäckens väggar. Slutligen återstår endast exocuticle och epicuticle av den gamla kutikulan, som underlagras av ny epicuticle och ny exocuticle. Djuret sväljer vatten, som det absorberar genom tarmen, och dess blodvolym ökar kraftigt. Det inre trycket får kutikulan att spricka och djuret drar sig ut ur sitt gamla exoskelett (figur 19-12). Därefter följer en sträckning av den fortfarande mjuka nya kutikulan, avlagring av den nya endokutikulan, omlagring av de räddade oorganiska salterna och andra beståndsdelar samt härdning av den nya kutikulan. Under perioden för skiftning är djuret försvarslöst och håller sig gömt.
 |
|
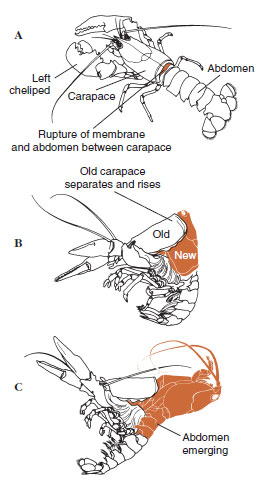 |
 |
| Figur 19-12 Förmultningssekvens hos en hummer, Homarus americanus. A, Membranen mellan carapace och buken spricker och carapace börjar långsamt höjas upp. Detta steg kan ta upp till 2 timmar. B och C: Huvudet, bröstkorgen och slutligen buken dras tillbaka. Denna process tar vanligtvis inte mer än 15 minuter. Omedelbart efter ecdysis är chelipederna uttorkade och kroppen är mycket mjuk. Hummern fortsätter att snabbt absorbera vatten så att kroppen inom 12 timmar ökar ca 20 % i längd och 50 % i vikt. Vävnadsvattnet kommer att ersättas av protein under de följande veckorna. |
Figur 19-11 Sekretion och resorption av hudceller vid ecdysis. |
När ett kräftdjur är ungt måste ecdysis inträffa ofta för att möjliggöra tillväxt, och multningscykeln är relativt kort. När djuret närmar sig mognad blir intermoltperioderna successivt längre, och hos vissa arter upphör moltningen helt och hållet. Under intermoltperioderna sker en ökning av vävnadsmassan när levande vävnad ersätter vatten.
Hormonell kontroll av ekdyscykeln
Och även om ekdysen är hormonellt kontrollerad initieras cykeln ofta av ett miljöstimulans som uppfattas av det centrala nervsystemet. Sådana stimuli kan vara temperatur, dagslängd och luftfuktighet (när det gäller landkrabbor). Signalen från det centrala nervsystemet minskar produktionen av ett hämmande hormon i X-organet. X-organet är en grupp neurosekretoriska celler i hjärnans medulla terminalis. Hos kräftor och andra decapoder finns medulla terminalis i ögonstammen. Hormonet transporteras i X-organets axoner till sinuskörteln (som i sig troligen inte har någon körtelfunktion), som också finns i ögonstjälken, där det frisätts i hemolymphen.
En sänkning av nivån av mullningshämmande hormon främjar frisättning av ett mullningshormon från Y-organen. Y-organen ligger under epidermis nära mandibelns adduktormuskler, och de är homol-ogus med prothorakala körtlar hos insekter, som producerar hormonet ecdyson. Multningshormonets verkan är att initiera processer som leder till ecdysis (proecdysis). När den väl har inletts fortsätter cykeln automatiskt utan ytterligare verkan av hormoner från vare sig X- eller Y-organen.
Andra endokrina funktioner: Det är inte bara så att avlägsnandet av ögonstjälkarna påskyndar ruggningen, man fann också för över 100 år sedan att kräftdjur vars ögonstjälkar har avlägsnats inte längre kan anpassa kroppens färgning till bakgrundsförhållandena. För över 50 år sedan upptäckte man att felet inte berodde på förlust av synen utan på förlust av hormoner i ögonstammarna. Kroppsfärgen hos kräftdjur är till stor del ett resultat av pigment i speciella förgrenade celler (kromatophorer) i epidermis.
Koncentration av pigmentkorn i mitten av cellerna ger en ljusare effekt, och spridning av pigment över hela cellerna ger en mörkare effekt. Pigmentbeteendet styrs av hormoner från neurosekretoriska celler i ögonstammen, liksom migrationen av retinapigment för ljus- och mörkeranpassning i ögonen (figur 19-8).
Utsläpp av neurosekretoriskt material från perikardialorganen i perikardväggarna orsakar en ökning av hjärtslagets frekvens och amplitud.
Androgena körtlar, som först hittades hos en amfipod (Orchestia, en vanlig strandhuggare), förekommer hos hanar av malakostracerna. Till skillnad från de flesta andra endokrina organ hos kräftdjur är dessa inte neurosekretoriska organ. Deras sekret stimulerar uttrycket av manliga könsegenskaper. Unga malacostracaner har rudimentära androgena körtlar, men hos honor utvecklas dessa körtlar inte. Om de implanteras artificiellt hos en hona förvandlas hennes äggstockar till testiklar och börjar producera spermier, och hennes bihang börjar få manliga kännetecken vid nästa skiftning. Hos isopoder finns de androgena körtlarna i testiklarna; hos alla andra malakostrakier finns de mellan musklerna i coxopoderna på de sista bröstbenen och är delvis fastsatta nära ändarna av vasa deferentia. Även om honorna inte har organ som liknar androgena körtlar producerar deras äggstockar ett eller två hormoner som påverkar de sekundära könsegenskaperna.
Hormoner som påverkar andra kroppsprocesser hos kräftdjur kan förekomma, och bevis tyder på att en neurosekretorisk substans som produceras i ögonstjälken reglerar blodsockernivån.
Föringsvanor
Föringsvanor och anpassningar för födosök varierar mycket bland kräftdjur. Många former kan växla från en typ av föda till en annan beroende på miljö och tillgång på föda, men alla använder samma grundläggande uppsättning munstycken. Mandibler och maxillae fungerar för att ta in föda; maxillipeds håller fast och krossar föda. Hos rovdjur tjänar de gående benen, särskilt chelipederna, till att fånga födan.
Många kräftdjur, både stora och små, är rovdjur, och vissa har intressanta anpassningar för att döda byten. En räkliknande form, Lygiosquilla, har på ett av sina gångben ett specialiserat finger som kan dras in i ett spår och släppas plötsligt för att genomborra ett förbipasserande byte. Pistolräkor (Alpheus spp.) har en enormt förstorad chela som kan spännas som hammaren på en pistol och knäppas med en kraft som bedövar bytet.
Suspensionsmänniskors föda sträcker sig från plankton och detritus till bakterier. Rovdjuren äter larver, maskar, kräftdjur, sniglar och fiskar. Ätare äter döda djur- och växtdelar. Suspensionsätare, t.ex. älvräkor, vattenloppor och havstulpaner, använder sina ben, som är försedda med en tjock frans av setae, för att skapa vattenströmmar som sveper med sig matpartiklar genom setae. Slamräkor (Upogebia spp.) använder långa setae på sina två första par bröstfästen för att sila matmaterial från vatten som cirkulerar genom deras hålor genom rörelser av simretterna.
Kräftor har en tvådelad mage (figur 19-13). Den första delen innehåller en magkvarn där föda, som redan rivits upp av mandiblerna, kan malas ytterligare av tre kalktänder till partiklar som är tillräckligt fina för att passera genom ett setosfilter i den andra delen; födapartiklarna passerar sedan in i tarmen för kemisk matsmältning.
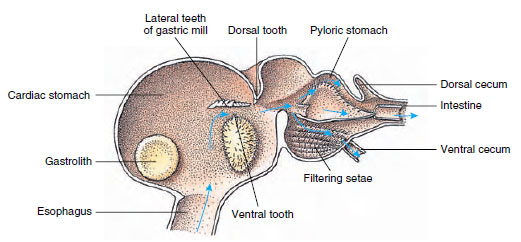
Figur 19-13
Malacostrakisk mage som visar gastrisk ”kvarn” och riktningar för matens rörelser. Kvarnen har kitinösa kammar, eller
tänder, för masticering, och setae för att sila maten innan den passerar in i pyloricmagsäcken.
Lämna ett svar