Liberação do neurotransmissor
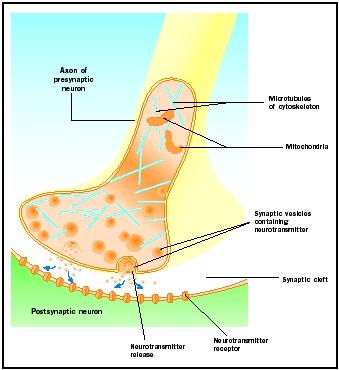
Potenciais de acção que chegam aos botões sinápticos activam a libertação do neurotransmissor para a fenda sináptica. O mecanismo molecular não é completamente compreendido. Um “atraso sináptico” de um a dois milissegundos ocorre entre a chegada do potencial de ação e a liberação do neurotransmissor. Os potenciais de ação abrem canais de cálcio na membrana do botão sináptico, o que causa um movimento interno de íons de cálcio . Os íons de cálcio provocam a liberação do neurotransmissor das vesículas sinápticas para a fenda sináptica. As vesículas sinápticas se fundem com a membrana pré-sináptica durante este processo de exocitose. As membranas das vesículas antigas tornam-se parte da membrana pré-sináptica e as novas vesículas se fixam a partir de uma área adjacente da membrana. Estas novas vesículas são posteriormente preenchidas com neurotransmissores recentemente sintetizados ou “reciclados”.
Os neurotransmissores liberados se difundem através da estreita fenda sináptica. Na membrana pós-sináptica, as moléculas neurotransmissoras ligam-se às moléculas receptoras ligadas à membrana com locais de reconhecimento específicos para aquele neurotransmissor. A ligação do neurotransmissor ao receptor desencadeia uma resposta pós-sináptica específica para esse receptor. Essas respostas podem ser excitatórias ou inibitórias, dependendo das propriedades do receptor. Se a estimulação do receptor resultar em que a membrana pós-sináptica se torne mais positiva eletricamente (despolarizada), é um potencial pós-sináptico excitatório (EPSP). Se mais negativo (hiperpolarizado), é um potencial pós-sináptico inibitório (IPSP). A excitação e inibição dependem das propriedades do receptor e não do neurotransmissor. Receptores acoplados a canais de sódio ou cálcio são excitatórios e produzem uma despolarização da membrana pós-sináptica, enquanto receptores acoplados a canais de cloreto ou potássio são inibidores e produzem uma hiperpolarização da membrana pós-sináptica. Tais receptores acoplados a canais de íons são chamados de receptores ionotrópicos.
Outros receptores são acoplados a sistemas de “segundo mensageiro” que iniciam uma série de reações bioquímicas na célula pós-sináptica. Estes são receptores metabotrópicos . Os receptores metabotrópicos podem produzir muitos diferentes
eventos pós-sinápticos. Estes variam desde a activação directa dos canais iónicos adjacentes, à alteração da sensibilidade do receptor, à transcrição de ácidos ribonucleicos mensageiros específicos (RNAs), ou mesmo à activação de genes específicos . As sinapses químicas fazem parte de um sistema de comunicação muito adaptável e flexível. Não são estruturas anatômicas estáticas com propriedades fixas, mas sim estruturas dinâmicas, capazes de alterar suas propriedades moleculares com a mudança das circunstâncias.
Existem literalmente centenas de neurotransmissores. Alguns são compostos bastante simples como a acetilcolina, a serotonina, as catecolaminas (dopamina, norepinefrina e epinefrina) e alguns dos aminoácidos. Muitos são mais complexos e pertencem à vasta gama de transmissores de neuropeptídeos. Uma vez liberados na fenda sináptica, os neurotransmissores permanecem ativos até que sejam alterados quimicamente ou levados de volta para o botão sináptico por sistemas de transporte especiais e reciclados. Nas sinapses colinérgicas, a acetilcolinesterase está presente na fenda sináptica. Esta enzima cliva o neurotransmissor em acetato e colina, nenhum dos quais está ativo. A serotonina e a epinefrina, por outro lado, são levadas para o terminal pré-sináptico e recicladas.
Deixe uma resposta