Subphylum Crustacea
General Nature of a Crustacea
Crustáceos diferem de outros artrópodes de várias maneiras, mas a única característica verdadeiramente distintiva é que os crustáceos são os únicos artrópodes com dois pares de antenas. Além de dois pares de antenas e um par de mandíbulas, os crustáceos possuem dois pares de maxilares na cabeça, seguidos por um par de apêndices em cada segmento do corpo ou somite. Em alguns crustáceos, nem todos os somitos têm apêndices. Todos os apêndices, exceto talvez as primeiras antenas, são primitivamente biramatosos (dois ramos principais), e pelo menos alguns apêndices de adultos de hoje mostram essa condição. Órgãos especializados em respiração, se presentes, funcionam como brânquias.
A maioria dos crustáceos tem entre 16 e 20 somites, mas algumas formas têm 60 somites ou mais. Um número maior de somitos é uma característica primitiva. A condição mais derivada é a de ter menos segmentos e maior tagmatização. Os principais tagmata são cabeça, tórax e abdômen, mas estes não são homólogos em toda a classe (ou mesmo dentro de algumas subclasses) devido aos vários graus de fusão dos somitos, por exemplo, como no cefalotórax.
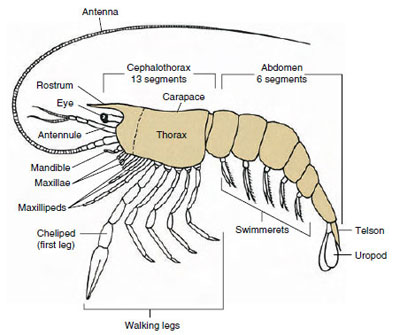
Figura 19-1
Plano arquetípico da Malacostraca. Os dois maxilares e três
maxilípedes foram separados diagramáticamente para ilustrar o plano
plano geral.
De longe o maior grupo de crustáceos é a classe Malacostraca, que inclui lagostas, caranguejos, camarões, tremonhas de praia, insetos de porcas, e muitos outros. Estes mostram uma disposição surpreendentemente constante de segmentos de corpo e tagmata, que é considerada o plano ancestral da classe (Figura 19-1). Este plano corporal típico tem uma cabeça de cinco (seis embrionariamente) somites fundidos, um tórax de oito somites e um abdômen de seis somites (sete em poucas espécies). Na extremidade anterior há uma tribuna não segmentada e na extremidade posterior há um telescópio não segmentado, que com o último somito abdominal e seus uropodes forma um ventilador de cauda em muitas formas.
Em muitos crustáceos a cutícula dorsal da cabeça pode se estender posteriormente e ao redor dos lados do animal para cobrir ou ser fundido com alguns ou todos os somitos torácicos e abdominais. Esta cobertura é chamada de carapaça. Em alguns grupos a carapaça forma válvulas tipo clamshell que cobrem a maior parte ou todo o corpo. Em decápodes (incluindo lagostas, camarões, caranguejos e outros), a carapaça cobre todo o cefalotórax mas não o abdômen.
Forma e Função
Por causa de seu tamanho e fácil disponibilidade, grandes crustáceos como os lagostim têm sido estudados mais do que outros grupos. Eles também são comumente estudados em cursos de laboratório introdutórios. Portanto, muitos dos comentários que se seguem aplicam-se especificamente aos lagostim e seus parentes.
Características Externas
Os corpos de crustáceos são cobertos com uma cutícula secretada composta de quitina, proteína e material calcário. As placas mais duras e pesadas de crustáceos maiores são particularmente altas em depósitos calcários. A cobertura protectora dura é macia e fina nas articulações entre os somitos, permitindo flexibilidade de movimento. A carapaça, se presente, cobre grande parte ou a totalidade do cefalotórax; em decápodes como os lagostim, todos os segmentos da cabeça e do tórax são fechados dorsalmente pela carapaça. Cada somito não fechado pela carapaça é coberto por uma placa cuticular dorsal, ou tergum (Figura 19-2A), e uma barra transversal ventral, o esterno, fica entre os apêndices segmentares (Figura 19-2B). O abdômen termina em um telescópio, que não é considerado um somito e carrega o ânus. (O telson pode ser homólogo ao pygidium annelidium.)
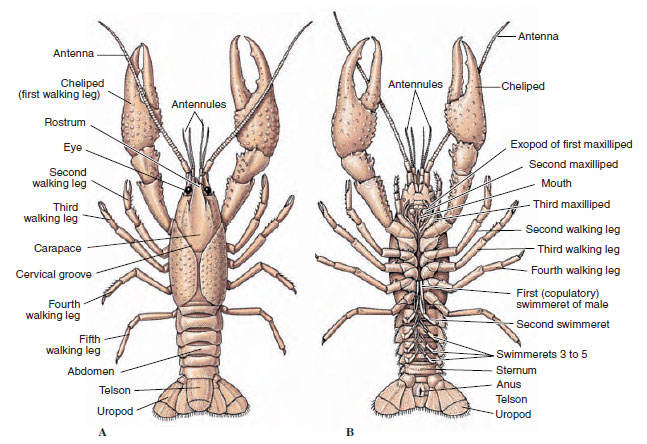
Figura 19-2
Estrutura externa de lagostim. A, Vista dorsal. B, Vista ventral.
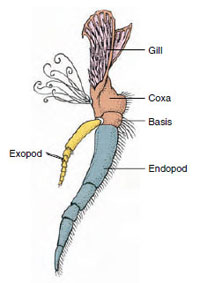
Figura 19-3
Partes de um crustáceo biramico
appendagem (terceiro maxilar de um
crayfish).
A posição dos gonoporos varia de acordo com o sexo e grupo de crustáceos. Eles podem estar sobre ou na base de um par de apêndices, na extremidade terminal do corpo, ou em somites sem pernas. Nos lagostins, as aberturas da vasa deferentia estão no lado mediano na base do quinto par de pernas andantes, e as dos oviductos estão na base do terceiro par. Nas fêmeas a abertura para o receptáculo seminal está geralmente localizada na linha mediana entre o quarto e o quinto par de pernas de caminhada.
Apendimentos: Os membros das classes Malacostraca (incluindo os lagostim) e Remipedia tipicamente têm um par de apêndices articulados em cada somito (Figura 19-3), embora os somitos abdominais das outras classes não tenham apêndices. Uma especialização considerável é evidente em apêndices de crustáceos derivados, como os lagostim. Entretanto, todas são variações do plano básico, biramous, ilustradas por um apêndice de lagostim como um maxiliped (um membro torácico modificado para tornar-se um apêndice de cabeça) (Figuras 19-3 e 19-4). A porção basal, ou protópode, tem um exópodo lateral e um endopépode medial. O protópode é composto por uma ou duas articulações (base e coxa), enquanto o exópodo e o endopépode têm de uma a várias articulações cada um. Alguns apêndices, como as pernas ambulantes de lagostim, são secundariamente pouco vistosos. Processos mediais ou laterais às vezes ocorrem em membros crustáceos, chamados endites e exits, respectivamente, e um exato no protópode é chamado epípoda. Os epípodas são frequentemente modificados como brânquias. A Tabela 19-1 mostra como os vários apêndices foram modificados a partir do plano biramatoso para se ajustarem a funções específicas.
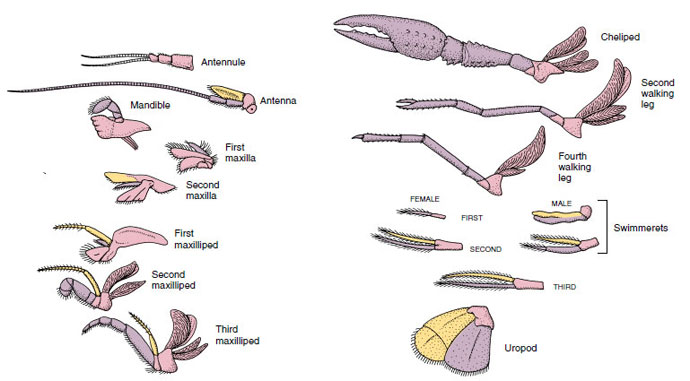
Figura 19-4
Aplicações de um lagostim mostrando como eles foram modificados a partir do plano biramônico básico, como encontrado em um nadador
. Protopod, rosa; endopod, roxo; exopod, amarelo.
Estruturas que têm um plano básico semelhante e desceram de uma forma comum são ditas homólogas, quer tenham a mesma função ou não. Uma vez que as pernas, partes da boca, quelípedos e nadadores especializados se desenvolveram a partir de um tipo biramônico comum, mas se modificaram para desempenhar funções diferentes, são todos homólogos entre si, uma condição conhecida como homologia em série. Primitivamente os membros eram todos muito semelhantes, mas durante a evolução das modificações estruturais, alguns ramos foram reduzidos, alguns perdidos, alguns muito alterados e algumas peças novas foram adicionadas. Os lagostim e seus aliados possuem a homologia serial mais elaborada do reino animal, tendo 17 tipos de apêndices distintos, mas serialmente homólogos (Tabela 19-1).
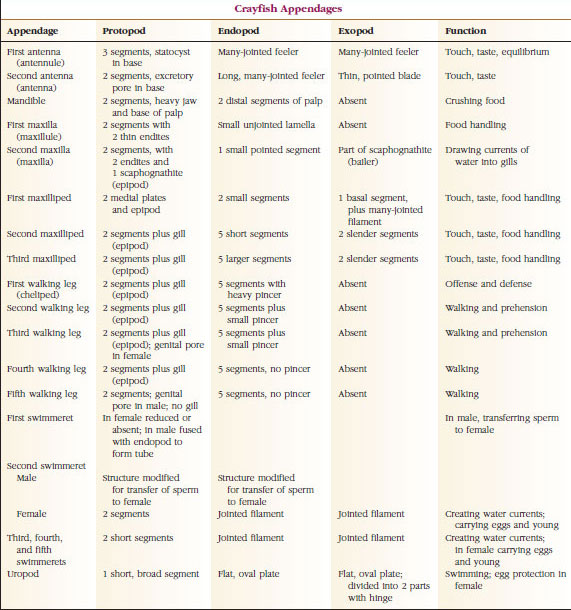
TABLE 19.1
Características internas
Os sistemas muscular e nervoso e a segmentação no tórax e abdômen mostram claramente metamerismo herdado de ancestrais anélidos, mas há modificações marcantes em outros sistemas. A maioria das modificações envolve concentração de partes em uma determinada região ou redução ou perda completa de partes, como a intersepta.
Hemocoel: O maior espaço corporal em artrópodes não é o celoma, mas um hemocoel cheio de sangue. Durante o desenvolvimento embrionário da maioria dos artrópodes, cavidades coelômicas vestigiais abrem-se dentro da mesoderme de pelo menos alguns somítas. Estas são logo obliteradas ou tornam-se contínuas com o espaço entre as estruturas mesodérmicas e ectodérmicas em desenvolvimento e a gema. Este espaço torna-se o hemocoel e, portanto, não é forrado por um peritônio mesodérmico. Nos crustáceos os únicos compartimentos coelômicos restantes são os sacos finais dos órgãos excretores e o espaço ao redor das gônadas.
Sistema Muscular: Os músculos estriados constituem uma parte considerável do corpo da maioria dos crustáceos. Os músculos são geralmente dispostos em grupos antagónicos: flexores, que atraem uma parte em direcção ao corpo, e extensores, que o estendem para fora. O abdómen de um lagostim tem poderosos flexores (Figura 19-5), que são usados quando o animal nada para trás – o melhor meio de escapar. Músculos fortes em ambos os lados do estômago controlam as mandíbulas.
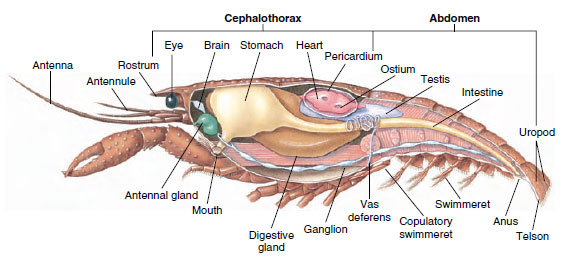
Figura 19-5
Estrutura interna de um lagostim macho.
Sistema respiratório: A troca de gases respiratórios em crustáceos menores ocorre em áreas mais finas da cutícula (por exemplo, nas pernas) ou em todo o corpo, e estruturas especializadas podem estar ausentes. Crustáceos maiores têm brânquias, que são delicadas, projeções em forma de penas com cutícula muito fina. Nos decápodes, os lados da carapaça envolvem a cavidade das brânquias, que é aberta anterior e ventralmente (Figura 19-6). As brânquias podem projectar-se da parede pleural para a cavidade das brânquias, da articulação das pernas torácicas com o corpo, ou da coxa torácica. Os dois últimos tipos são típicos dos lagostins de cera. O “bailer”, uma parte da segunda maxila, atrai água sobre os filamentos das brânquias, para dentro da cavidade das brânquias nas bases das pernas, e para fora da cavidade das brânquias na parte anterior.
Sistema Circulatório: Os crustáceos e outros artrópodes têm um tipo de sistema circulatório “aberto” ou lacunar. Isto significa que não existem veias nem separação do sangue do líquido intersticial, como acontece nos animais com sistemas fechados. A hemolinfa (sangue) deixa o coração através das artérias, circula através do hemocoel e retorna aos seios venosos, ou espaços, em vez de veias, antes de reentrar no coração. Os anelídeos têm um sistema fechado, assim como os vertebrados.
Um coração dorsal é o principal órgão propulsor. É um saco único de músculo estriado. A hemolinfa entra no coração do seio pericárdico circundante através dos óstios pareados, com válvulas que impedem o refluxo para dentro do seio (Figura 19-6). A hemolinfa do coração entra em uma ou mais artérias. As válvulas nas artérias impedem o refluxo da hemolinfa. Pequenas artérias esvaziam-se nos seios torácicos, que por sua vez descarregam frequentemente para um grande seio esterno (Figura 19-6).
De lá, os canais sinusais aferentes transportam a hemolinfa para as guelras, se presentes, para a troca de oxigénio e dióxido de carbono. A hemolinfa então retorna ao seio pericárdico pelos canais eferentes (Figura 19-6).
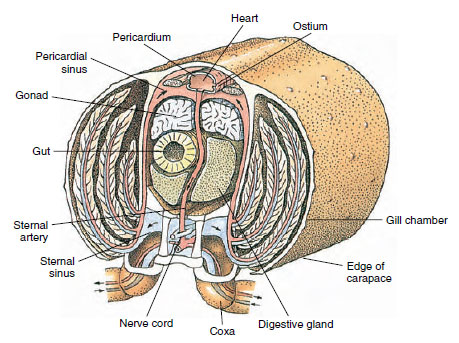
Figura 19-6
secção transversal diagramática através da região cardíaca de um lagostim mostrando a direcção do fluxo sanguíneo neste sistema “aberto”
sangue. O coração bombeia o sangue para os tecidos do corpo através das artérias, que se esvaziam nos seios dos tecidos. Retornando
sangue entra no seio esterno, depois passa pelas guelras para troca de gases, e finalmente de volta ao seio pericárdico por eferentes
canais. Note a ausência de veias.
Hemolinfa em artrópodes é em grande parte incolor. Inclui células ameboidais de pelo menos dois tipos. Hemocianina, um pigmento respiratório contendo cobre, ou hemoglobina, um pigmento contendo ferro, pode ser transportado em solução. A hemolinfa tem a propriedade de coagular, o que evita a sua perda em lesões menores. Algumas células ameboidais liberam um coagulante tipo trombina que precipita a coagulação.
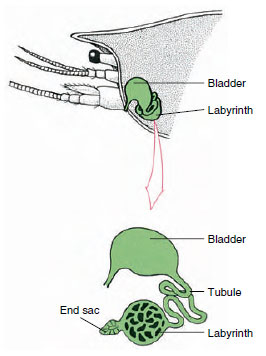
Figura 19-7
Esquema de glândula antenal (glândula verde) de
crayfishes. (Em posição natural o órgão é muito
dobrado.) Alguns crustáceos não possuem um labirinto,
e o tubo excretor (canal nefróide) é
um tubo muito enrolado.
>
Sistema excretor: Os órgãos excretores de crustáceos adultos são um par de estruturas tubulares localizadas na parte ventral da cabeça anterior ao esôfago (Figura 19-5). Eles são chamados de glândulas antenais ou glândulas maxilares, dependendo se abrem na base das antenas ou da segunda maxila. Alguns crustáceos adultos têm ambos. Os órgãos excretores dos decápodes são glândulas antenais, também chamadas de glândulas verdes neste grupo. Os crustáceos não possuem túbulos malpighianos, os órgãos excretores de aranhas e insetos.
O saco final da glândula antenal, que é derivado de um compartimento celômico embrionário, consiste de uma pequena vesícula (bolsa) e uma massa esponjosa chamada labirinto. O labirinto liga-se por um túbulo excretor a uma bexiga dorsal, que se abre para o exterior por um poro na superfície ventral do segmento basal da antena (Figura 19-7). A pressão hidrostática dentro do hemocoel fornece força para a filtração do líquido para o saco final. Como o filtrado passa pelo tubo excretor e pela bexiga, ele é modificado pela reabsorção de sais, aminoácidos, glicose e alguma água e finalmente é excretado como urina.
A excreção de resíduos nitrogenados (principalmente amônia) ocorre por difusão através de áreas finas da cutícula, especialmente as brânquias, e os chamados órgãos excretores funcionam principalmente para regular a composição iônica e osmótica dos fluidos corporais. Os crustáceos de água doce, como os lagostim, são constantemente ameaçados de diluição excessiva do seu sangue pela água, que se difunde através das guelras e outras superfícies permeáveis à água. As glândulas verdes, ao formar uma urina diluída e pouco salgada, actuam como um eficaz dispositivo de “controlo de inundação”. Alguns Na+ e Cl- são perdidos na urina, mas esta perda é compensada pela absorção activa do sal dissolvido pelas brânquias. Em crustáceos marinhos, como lagostas e caranguejos, o rim funciona para ajustar a composição salina da hemolinfa pela modificação seletiva do conteúdo de sal da urina tubular. Nestas formas, a urina permanece isosmótica ao sangue.
Sistemas nervosos e sensoriais: Os sistemas nervosos de crustáceos e anelídeos têm muito em comum, embora os dos crustáceos tenham mais fusão de gânglios (Figura 19-5). O cérebro é um par de gânglios supra-esofágicos que fornece nervos aos olhos e dois pares de antenas. Ele é unido por conectivos ao gânglio subesofágico, uma fusão de pelo menos cinco pares de gânglios que fornecem nervos para a boca, apêndices, esôfago e glândulas antenais. O cordão nervoso ventral duplo tem um par de gânglios para cada somito e nervos que servem os apêndices, músculos e outras partes.
Além deste sistema central, pode haver um sistema nervoso simpático associado ao trato digestivo.
Os crustáceos têm órgãos sensoriais melhor desenvolvidos do que os anelídeos. Os maiores órgãos sensoriais dos lagostins são os olhos e os estatocistos. Amplamente distribuídos pelo corpo são pêlos táteis, delicadas projeções de cutícula que são especialmente abundantes em quelônios, partes da boca e telescópios. Os sentidos químicos do gosto e do olfato são encontrados nos pêlos das antenas, partes da boca e outros lugares.
Um estatocisto semelhante a um saco, aberto para a superfície por um poro dorsal, é encontrado no segmento basal de cada primeira antena de lagostim. O estatocisto contém uma crista que contém pêlos sensoriais formados a partir do revestimento quitinoso e grãos de areia que servem de estatólitos. Sempre que o animal muda de posição, mudanças correspondentes na posição dos grãos nos pêlos sensoriais são transmitidas como estímulo ao cérebro, e o animal pode se ajustar de acordo. Cada molt (ecdise) de cutícula resulta na perda do revestimento cuticular do estatocisto e com ele os grãos de areia. Novos grãos são coletados através do poro dorsal após a ecdise.
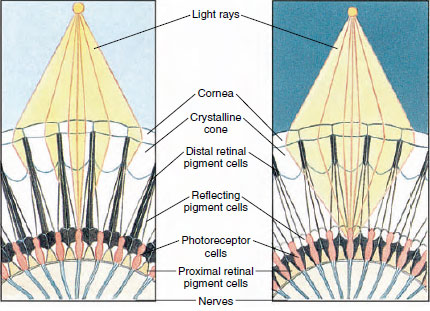
Figura 19-8
Porção de olho composto de um artrópode mostrando migração de pigmento
em ommatidia para visão diurna e noturna. Cinco ommatidia são representados em
cada diagrama. Durante o dia, cada ommatidium é rodeado por um colar de pigmento escuro
de modo que cada ommatidium é estimulado apenas pelos raios de luz
que entram na sua própria córnea (visão em mosaico); durante a noite, os pigmentos formam
colares completos e os raios de luz podem espalhar-se para ommatidia adjacente
(contínua, ou sobreposição, imagem).
Os olhos em muitos crustáceos são compostos, compostos de muitas unidades fotorreceptoras chamadas ommatidia (Figura 19-8). Cobrindo a superfície arredondada de cada olho está uma área transparente da cutícula, a córnea, que é dividida em muitos pequenos quadrados ou hexágonos conhecidos como facetas. Estas facetas são as extremidades externas da ommatidia. Cada ommatidium comporta-se como um olho minúsculo e contém vários tipos de células dispostas de forma colunar (Figura 19-8). Células de pigmento preto são encontradas entre ommatidia adjacente.
O movimento do pigmento em um olho composto de artrópodes permite que ele se ajuste para diferentes quantidades de luz. Existem três conjuntos de células de pigmento em cada ommatidium: retina distal, retina proximal e reflectora; estes estão dispostos de tal forma que podem formar um colar ou manga mais ou menos completa ao redor de cada ommatidium. Para uma forte adaptação à luz ou ao dia, o pigmento da retina distal move-se para dentro e encontra o pigmento da retina proximal que se move para fora, de modo a formar uma manga de pigmento completa ao redor do ommatidium (Figura 19-8). Nesta condição, apenas os raios que atingem diretamente a córnea alcançarão as células fotorreceptoras (retinulares), pois cada ommatidium está protegido de outros. Assim, cada ommatidíaco verá apenas uma área limitada do campo de visão (um mosaico, ou imagem de aposição). Na luz fraca os pigmentos distal e proximal separam-se para que os raios de luz, com o auxílio de células de pigmento refletor, tenham a chance de se espalhar para ommatidia adjacente e formar uma imagem contínua, ou sobreposição. Este segundo tipo de visão é menos precisa mas aproveita ao máximo a quantidade limitada de luz recebida.
Reprodução, Ciclos de Vida e Função Endócrina
A maioria dos crustáceos tem sexos separados, e há várias especializações para a cópula entre diferentes grupos. As cracas são monóicas, mas geralmente praticam a fertilização cruzada. Em alguns ostracodes os machos são escassos, e a reprodução é geralmente partenogenética. A maioria dos crustáceos cria os seus ovos de alguma forma: os ramos e as cracas têm câmaras de criação especiais, os copépodes têm sacos de criação presos aos lados do abdómen (ver figura 19-19) e muitos malacostracanos transportam ovos e crias presas aos seus apêndices abdominais.
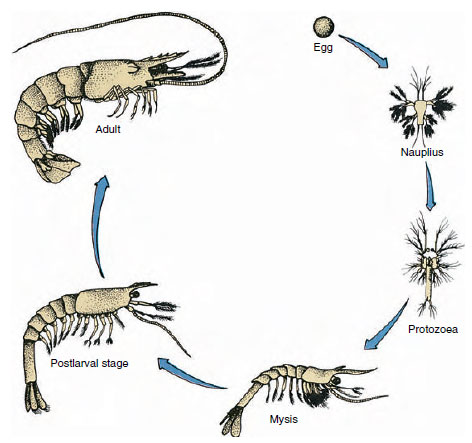
Figura 19-9
Ciclo de vida de um Penaeus camarão do Golfo do México. Os penaeus desovam em profundidades de 40 a 90 m. As formas larvares jovens são
planctónicas e deslocam-se para terra para águas de menor salinidade para se desenvolverem como juvenis. Os camarões mais velhos regressam a águas mais profundas
offshore.
Crayfishes têm desenvolvimento directo: não há forma larval. Um pequeno juvenil com a mesma forma que o adulto e um conjunto completo de apêndices e somitos que eclodem do ovo. Contudo, o desenvolvimento é indirecto na maioria dos crustáceos, e uma larva muito diferente do adulto na estrutura e aparência eclodem do ovo. A mudança da larva para um adulto é, em última análise, uma metamorfose. A larva primitiva e a mais comum nos crustáceos é o nauplius (Figura 19-9 e 19-23). Os nauplius têm apenas três pares de apêndices: primeiras antenas uniramas, antenas biramas e mandíbulas biramas. Todos funcionam como apêndices de natação nesta fase.
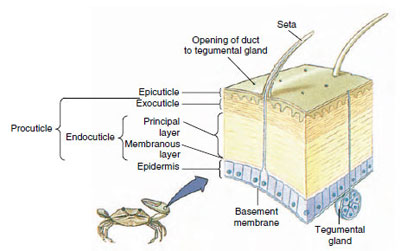
Figura 19-10
Estrutura da cutícula do crustáceo.
Desenvolvimento subseqüente pode envolver uma mudança gradual na forma do corpo adulto, e apêndices e somites são adicionados através de uma série de moldes, ou a suposição da forma adulta pode envolver mudanças mais abruptas. Por exemplo, a metamorfose de um craca procede de um nauplius nadador livre para uma larva com uma carapaça bivalve chamada cipride e finalmente para um adulto sésseis com placas calcárias.
Ecdise: A ecdise (ek’duh-sis) (Gr. ekdyein, para se despir), ou moldagem, é necessária para que o corpo aumente de tamanho porque o exoesqueleto não é vivo e não cresce conforme o animal cresce. Muito do funcionamento de um crustáceo, incluindo sua reprodução, comportamento e muitos processos metabólicos, é diretamente afetado pela fisiologia do ciclo de moldagem.
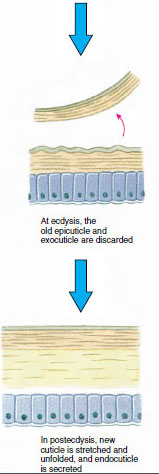
Cuticle, que é secretado pela epiderme subjacente, tem várias camadas (Figura 19-10). A mais externa é a epicúla, uma camada muito fina de proteína impregnada de lipidimpregnação. A maior parte da cutícula é constituída por várias camadas de procuícula: (1) exocuícula, que está logo abaixo da epicutícula e contém proteínas, sais de cálcio e quitina; (2) endocutícula, que é composta por (3) uma camada principal, que contém mais quitina e menos proteína e é fortemente calcificada, e (4) uma camada membranosa não calcificada, uma camada relativamente fina de quitina e proteína.
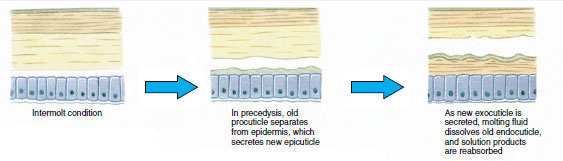
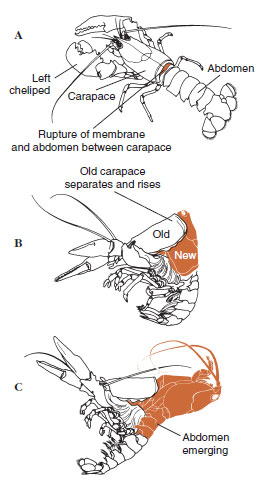
Algum tempo antes da ecdise propriamente dita, as células epidérmicas aumentam consideravelmente. Elas se separam da camada membranosa, secretam uma nova epicutícula e começam a secretar um novo exocuticulo (Figura 19-11). Enzimas são liberadas na área acima da nova epicutícula. Estas enzimas começam a dissolver a antiga endocutícula, e os produtos solúveis são reabsorvidos e armazenados dentro do corpo do crustáceo. Alguns sais de cálcio são armazenados como gastrólitos (acreções minerais) nas paredes do estômago. Finalmente, só restam exocuticula e epicuticula da cutícula antiga, subjacente a nova epicuticula e a nova exocuticula. O animal engole água, que absorve através do intestino, e o seu volume sanguíneo aumenta muito. A pressão interna faz com que a cutícula se parta, e o animal se puxa para fora do seu antigo exosqueleto (Figura 19-12). Segue-se então um alongamento da nova cutícula ainda mole, deposição da nova endocutícula, redefinição dos sais inorgânicos recuperados e outros constituintes, e endurecimento da nova cutícula. Durante o período de moldagem, o animal está indefeso e permanece escondido.
 |
|
 |
 |
| Figura 19-12 Sequência de polimento em uma lagosta, Homarus americanus. A, Membrana entre carapaça e rupturas do abdômen, e a carapaça começa a elevação lenta. Este passo pode demorar até 2 horas. B e C, cabeça, tórax, e finalmente o abdômen se retira. Este processo geralmente não leva mais do que 15 minutos. Imediatamente após a ecdise, os quelípedos são dessecados e o corpo é muito macio. A lagosta continua a absorver rapidamente a água de modo que dentro de 12 horas o corpo aumenta cerca de 20% em comprimento e 50% em peso. A água dos tecidos será substituída por proteína nas semanas seguintes. |
Figura 19-11 Secreção de partículas e resorção em ecdise. |
Quando um crustáceo é jovem, a ecdise deve ocorrer frequentemente para permitir o crescimento, e o ciclo de moldagem é relativamente curto. À medida que o animal se aproxima da maturidade, os períodos de intermoltagem tornam-se progressivamente mais longos e, em algumas espécies, a fusão cessa por completo. Durante os períodos de intermolt, o aumento da massa tecidual ocorre à medida que o tecido vivo substitui a água.
Controle hormonal do ciclo de ecdise
Embora a ecdise seja controlada hormonalmente, o ciclo é frequentemente iniciado por um estímulo ambiental percebido pelo sistema nervoso central. Tais estímulos podem incluir temperatura, duração do dia e umidade (no caso de caranguejos terrestres). O sinal do sistema nervoso central diminui a produção de uma hormona inibidora de moléstias pelo órgão X. O órgão X é um grupo de células neurosecretoras na medula terminal do cérebro. Em lagostim e outros decápodes, a medula terminal é encontrada na base do olho. O hormônio é carregado nos axônios do órgão X até a glândula sinusal (que provavelmente não é glandular em função), também na base do olho, onde é liberado na hemolinfa.
Uma queda no nível de hormônio inibidor de moltamento promove a liberação de um hormônio de moltamento dos Y-organs. Os Yorgans estão abaixo da epiderme perto dos músculos adutores das mandíbulas, e são homol-ogus para as glândulas protorácicas dos insetos, que produzem a hormona ecdisona. A acção da hormona de moldagem é iniciar processos que levam à ecdise (proecdise). Uma vez iniciado, o ciclo prossegue automaticamente sem a ação adicional de hormônios do X- ou Y-organs.
Outras Funções Endócrinas: Não só a remoção dos apoios oculares acelera a moldagem, como também foi descoberto há mais de 100 anos que os crustáceos cujos apoios oculares foram removidos já não conseguem ajustar a coloração do corpo às condições de fundo. Há mais de 50 anos foi descoberto que o defeito não era causado pela perda da visão, mas pela perda de hormônios nos apoios oculares. A cor corporal dos crustáceos é em grande parte resultado de pigmentos em células ramificadas especiais (cromatóforos) na epiderme.
Concentração dos grânulos de pigmento no centro das células causa um efeito de clareamento, e a dispersão do pigmento por todas as células causa um efeito de escurecimento. O comportamento dos pigmentos é controlado por hormônios das células neurosecretorianas na base ocular, assim como a migração do pigmento retiniano para adaptação à luz e à escuridão nos olhos (Figura 19-8).
O descolamento do material neurosecretoriano dos órgãos pericárdicos na parede do pericárdio causa um aumento na frequência e amplitude do batimento cardíaco.
Glândulas androgênicas, encontradas primeiramente em um anfípoda (Orchestia, uma tremonha de praia comum), ocorrem em malacostracanos masculinos. Ao contrário da maioria dos outros órgãos endócrinos em crustáceos, estes não são órgãos neurosecretos. A sua secreção estimula a expressão das características sexuais masculinas. Os jovens malacostracanos possuem glândulas rudimentares androgênicas, mas nas fêmeas essas glândulas não se desenvolvem. Se são implantadas artificialmente numa fêmea, os seus ovários transformam-se em testículos e começam a produzir esperma, e os seus apêndices começam a assumir características masculinas na muda seguinte. Nos isópodos as glândulas androgênicas são encontradas nos testículos; em todos os outros malacostracanos estão entre os músculos dos coxópodes das últimas pernas torácicas e parcialmente presas perto das extremidades da vasa deferentia. Embora as fêmeas não possuam órgãos semelhantes às glândulas androgênicas, seus ovários produzem um ou dois hormônios que influenciam as características sexuais secundárias.
Hormônios que influenciam outros processos corporais em crustáceos podem estar presentes, e evidências sugerem que uma substância neurosecretora produzida na base ocular regula o nível de açúcar no sangue.
Hábitos alimentares
Hábitos alimentares e adaptações para alimentação variam muito entre os crustáceos. Muitas formas podem mudar de um tipo de alimentação para outro dependendo do ambiente e da disponibilidade de alimentos, mas todas utilizam o mesmo conjunto fundamental de partes da boca. As mandíbulas e maxilares funcionam para ingerir alimentos; os maxilípedes seguram e esmagam os alimentos. Nos predadores as pernas andantes, particularmente os quelípedos, servem na captura de alimentos.
Muitos crustáceos, grandes e pequenos, são predadores, e alguns têm adaptações interessantes para matar as presas. Uma forma de shrimplike, Lygiosquilla, tem em uma de suas patas andantes um dígito especializado que pode ser arrastado para uma ranhura e liberado subitamente para perfurar as presas que passam. Os camarões pistola (Alpheus spp.) têm um chela enormemente aumentado que pode ser armado como o martelo de uma arma e quebrado com uma força que atordoa suas presas.
O alimento dos alimentadores de suspensão varia de plâncton e detritos a bactérias. Os predadores consomem larvas, minhocas, crustáceos, caracóis e peixes. Os necrófagos comem matéria vegetal e animal morta. Alimentadores de suspensão, como os camarões-fadas, pulgas d’água e cracas, usam suas patas, que carregam uma grossa franja de sedas, para criar correntes de água que varrem partículas de alimento pelas sedas. Os camarões de lama (Upogebia spp.) usam sedas longas em seus dois primeiros pares de apêndices torácicos para esticar o material de alimento da água que circula através de sua toca pelos movimentos de seus nadadores.
Os lagostins têm um estômago de duas partes (Figura 19-13). A primeira parte contém um moinho gástrico no qual os alimentos, já rasgados pelas mandíbulas, podem ser moídos por três dentes calcários em partículas suficientemente finas para passar por um filtro de setose na segunda parte; as partículas dos alimentos passam então para o intestino para digestão química.
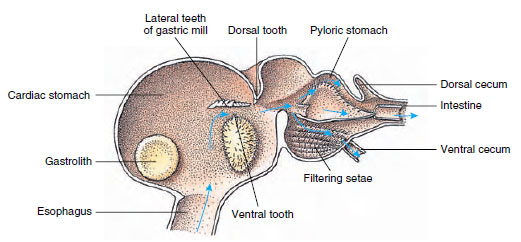
Figura 19-13
Estômago do malacostracan mostrando o “moinho” gástrico e as direções dos movimentos dos alimentos. Moinho tem cristas quitínicas, ou
teeth, para mastigação, e sedas para coar os alimentos antes de passar para o estômago pilórico.
Deixe uma resposta