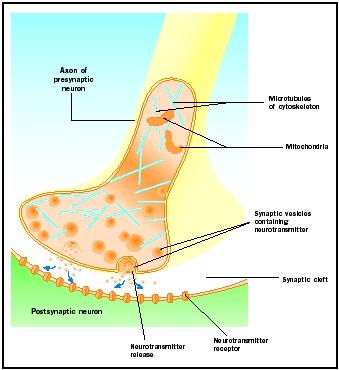
Libération de neurotransmetteur
Les potentiels d’action arrivant aux boutons synaptiques déclenchent la libération de neurotransmetteur dans la fente synaptique. Le mécanisme moléculaire n’est pas complètement compris. Un « délai synaptique » d’une à deux millisecondes se produit entre l’arrivée du potentiel d’action et la libération du neurotransmetteur. Les potentiels d’action ouvrent des canaux calciques dans la membrane du bouton synaptique, ce qui provoque un mouvement vers l’intérieur des ions calcium . Les ions calcium déclenchent la libération du neurotransmetteur des vésicules synaptiques dans la fente synaptique. Les vésicules synaptiques fusionnent avec la membrane présynaptique au cours de ce processus d’exocytose. Les membranes des anciennes vésicules deviennent une partie de la membrane présynaptique et de nouvelles vésicules se détachent d’une zone adjacente de la membrane. Ces nouvelles vésicules sont ensuite remplies à nouveau de neurotransmetteurs nouvellement synthétisés ou « recyclés » .
Les neurotransmetteurs libérés diffusent à travers l’étroite fente synaptique. Au niveau de la membrane postsynaptique, les molécules de neurotransmetteur se lient à des molécules réceptrices liées à la membrane et dotées de sites de reconnaissance spécifiques à ce neurotransmetteur. La liaison du neurotransmetteur au récepteur déclenche une réponse postsynaptique spécifique à ce récepteur. Ces réponses peuvent être soit excitatrices, soit inhibitrices, en fonction des propriétés du récepteur. Si la stimulation du récepteur a pour effet de rendre la membrane postsynaptique plus positive électriquement (dépolarisée), il s’agit d’un potentiel postsynaptique excitateur (EPSP). Si elle est plus négative (hyperpolarisée), il s’agit d’un potentiel postsynaptique inhibiteur (IPSP). L’excitation et l’inhibition dépendent des propriétés du récepteur et non du neurotransmetteur. Les récepteurs couplés à des canaux sodiques ou calciques sont excitateurs et produisent une dépolarisation de la membrane postsynaptique, tandis que les récepteurs couplés à des canaux chlorure ou potassium sont inhibiteurs et produisent une hyperpolarisation de la membrane postsynaptique. De tels récepteurs couplés à des canaux ioniques sont appelés récepteurs ionotropes.
D’autres récepteurs sont couplés à des systèmes de « second messager » qui initient une série de réactions biochimiques dans la cellule postsynaptique. Ce sont les récepteurs métabotropes . Les récepteurs métabotropes peuvent produire de nombreux
événements postsynaptiques. Ceux-ci vont de l’activation directe des canaux ioniques adjacents, à la modification de la sensibilité des récepteurs, à la transcription d’acides ribonucléiques messagers (ARN) spécifiques, voire à l’activation de gènes spécifiques . Les synapses chimiques font partie d’un système de communication très adaptable et flexible. Ce ne sont pas des structures anatomiques statiques aux propriétés fixes, mais des structures dynamiques, capables de modifier leurs propriétés moléculaires en fonction des circonstances.
Il existe littéralement des centaines de neurotransmetteurs. Certains sont des composés assez simples comme l’acétylcholine, la sérotonine, les catécholamines (dopamine, norépinéphrine et épinéphrine) et un certain nombre d’acides aminés . Beaucoup sont plus complexes et font partie de la vaste gamme de neuropeptides transmetteurs. Une fois libérés dans la fente synaptique, les neurotransmetteurs restent actifs jusqu’à ce qu’ils soient soit modifiés chimiquement, soit ramenés dans le bouton synaptique par des systèmes de transport spéciaux et recyclés. Au niveau des synapses cholinergiques, l’acétylcholinestérase est présente dans la fente synaptique. Cette enzyme clive le neurotransmetteur en acétate et en choline, qui ne sont pas actifs. La sérotonine et l’épinéphrine, en revanche, sont absorbées dans le terminal présynaptique et recyclées.
Laisser un commentaire