Sous-embranchement des crustacés
Nature générale d’un crustacé
Les crustacés diffèrent des autres arthropodes de diverses manières, mais la seule caractéristique vraiment distinctive est que les crustacés sont les seuls arthropodes à posséder deux paires d’antennes. En plus de deux paires d’antennes et d’une paire de mandibules, les crustacés possèdent deux paires de maxillaires sur la tête, suivies d’une paire d’appendices sur chaque segment du corps ou somite. Chez certains crustacés, tous les somites ne portent pas d’appendices. Tous les appendices, à l’exception peut-être des premières antennes, sont primitivement biramus (deux branches principales), et au moins certains appendices des adultes actuels présentent cette condition. Les organes spécialisés pour la respiration, s’ils sont présents, fonctionnent comme des branchies.
La plupart des crustacés ont entre 16 et 20 somites, mais certaines formes ont 60 somites ou plus. Un plus grand nombre de somites est une caractéristique primitive. La condition plus dérivée est d’avoir moins de segments et une tagmatisation accrue. Les principaux tagmates sont la tête, le thorax et l’abdomen, mais ceux-ci ne sont pas homologues dans toute la classe (ou même dans certaines sous-classes) en raison de degrés variables de fusion des somites, par exemple, comme dans le céphalothorax.
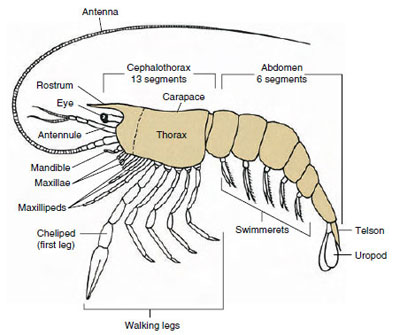
Figure 19-1
Plan archétypique de Malacostraca. Les deux maxillaires et les trois
maxillipèdes ont été séparés schématiquement pour illustrer le
plan général.
Le groupe de crustacés de loin le plus important est la classe des Malacostraca, qui comprend les homards, les crabes, les crevettes, les sauterelles de plage, les cloportes et bien d’autres. Ceux-ci présentent une disposition étonnamment constante des segments corporels et des tagmata, qui est considérée comme le plan ancestral de la classe (figure 19-1). Ce plan corporel typique comporte une tête de cinq somites fusionnés (six au stade embryonnaire), un thorax de huit somites et un abdomen de six somites (sept chez quelques espèces). A l’extrémité antérieure se trouve un rostre non segmenté et à l’extrémité postérieure un telson non segmenté qui, avec le dernier somite abdominal et ses uropodes, forme un éventail caudal dans de nombreuses formes.
Dans de nombreux crustacés, la cuticule dorsale de la tête peut s’étendre vers l’arrière et autour des côtés de l’animal pour couvrir ou être fusionnée avec une partie ou la totalité des somites thoraciques et abdominaux. Cette couverture s’appelle une carapace. Chez certains groupes, la carapace forme des valves en forme de coquille qui recouvrent la plupart ou la totalité du corps. Chez les décapodes (y compris les homards, les crevettes, les crabes et autres), la carapace couvre tout le céphalothorax mais pas l’abdomen.
Forme et fonction
En raison de leur taille et de leur facilité d’accès, les grands crustacés comme les écrevisses ont été étudiés plus que les autres groupes. Ils sont également couramment étudiés dans les cours d’introduction au laboratoire. Par conséquent, de nombreux commentaires qui suivent s’appliquent spécifiquement aux écrevisses et à leurs proches.
Marques externes
Le corps des crustacés est recouvert d’une cuticule sécrétée composée de chitine, de protéines et de matière calcaire. Les plaques plus dures et lourdes des grands crustacés sont particulièrement riches en dépôts calcaires. Le revêtement protecteur dur est souple et fin au niveau des articulations entre les somites, ce qui permet la flexibilité des mouvements. La carapace, si elle est présente, couvre une grande partie ou la totalité du céphalothorax ; chez les décapodes comme les écrevisses, tous les segments de la tête et du thorax sont enfermés dorsalement par la carapace. Chaque somite non entouré par la carapace est recouvert d’une plaque cuticulaire dorsale, ou tergum (Figure 19-2A), et une barre transversale ventrale, le sternum, se trouve entre les appendices segmentaires (Figure 19-2B). L’abdomen se termine par un telson, qui n’est pas considéré comme un somite et qui porte l’anus. (Le telson peut être homologue au pygidium des annélides.)
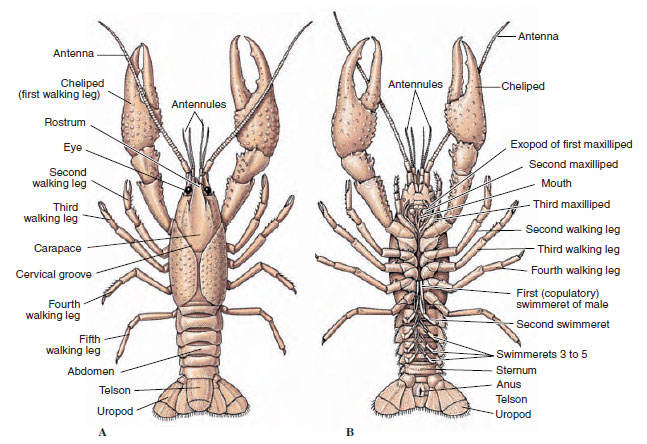
Figure 19-2
Structure externe des écrevisses. A, vue dorsale. B, vue ventrale.
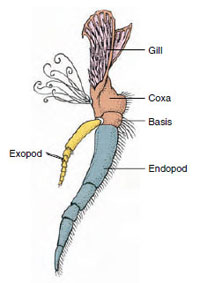
Figure 19-3
Parties d’un
appendice de crustacé biramique (troisième maxillipède d’une
écrevisse).
La position des gonopores varie selon le sexe et le groupe de crustacés. Ils peuvent être sur ou à la base d’une paire d’appendices, à l’extrémité terminale du corps, ou sur des somites sans pattes. Chez les écrevisses, les ouvertures des canaux déférents se trouvent sur le côté médian à la base de la cinquième paire de pattes ambulatoires, et celles des oviductes à la base de la troisième paire. Chez les femelles, l’ouverture du réceptacle séminal est généralement située sur la ligne médiane ventrale entre les quatrième et cinquième paires de pattes de marche.
Appendices : Les membres des classes Malacostraca (y compris les écrevisses) et Remipedia possèdent généralement une paire d’appendices articulés sur chaque somite (figure 19-3), bien que les somites abdominaux des autres classes ne portent pas d’appendices. Une spécialisation considérable est évidente dans les appendices des crustacés dérivés tels que les écrevisses. Cependant, tous sont des variations du plan de base biramus, illustré par un appendice d’écrevisse comme le maxillipède (un membre thoracique modifié pour devenir un appendice de la tête) (Figures 19-3 et 19-4). La partie basale, ou protopode, porte un exopode latéral et un endopode médian. Le protopode est constitué d’une ou deux articulations (base et coxa), alors que l’exopode et l’endopode ont chacun de une à plusieurs articulations. Certains appendices, comme les pattes marcheuses des écrevisses, sont devenus secondairement uniramides. Les membres des crustacés présentent parfois des processus médians ou latéraux, appelés respectivement endites et exites, et un exite sur le protopode est appelé un épipode. Les épipodes sont souvent modifiés en branchies. Le tableau 19-1 montre comment les différents appendices se sont modifiés à partir du plan biramus pour répondre à des fonctions spécifiques.
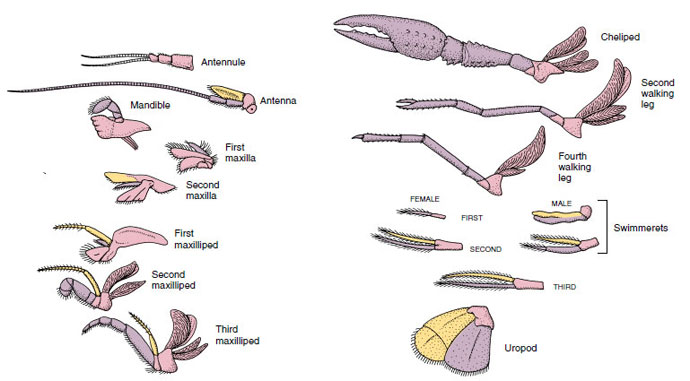
Figure 19-4
Appendices d’une écrevisse montrant comment ils se sont modifiés à partir du plan biramique de base, comme on le trouve chez un
swimmeret. Protopode, rose ; endopode, violet ; exopode, jaune.
Les structures qui ont un plan de base similaire et qui sont descendues d’une forme commune sont dites homologues, qu’elles aient ou non la même fonction. Puisque les pattes de marche spécialisées, les pièces buccales, les chélipèdes et les nageoires se sont tous développés à partir d’un type biramique commun, mais se sont modifiés pour remplir des fonctions différentes, ils sont tous homologues les uns des autres, une condition connue sous le nom d’homologie sérielle. Primitivement, les membres étaient tous très semblables, mais au cours de l’évolution des modifications structurelles, certaines branches ont été réduites, d’autres perdues, d’autres encore fortement modifiées, et de nouvelles parties ajoutées. Les écrevisses et leurs alliés possèdent l’homologie sérielle la plus élaborée du règne animal, avec 17 types d’appendices distincts mais homologues en série (tableau 19-1).
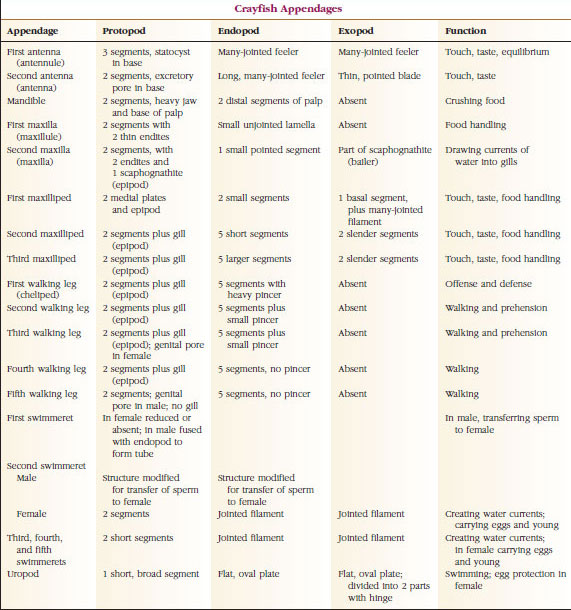
TABLE 19.1
Caractéristiques internes
Les systèmes musculaire et nerveux et la segmentation du thorax et de l’abdomen montrent clairement un métamérisme hérité d’ancêtres de type annélide, mais il existe des modifications marquées dans d’autres systèmes. La plupart des changements impliquent la concentration de parties dans une région particulière ou bien la réduction ou la perte complète de parties, comme l’intersepta.
Hémocoel : Le principal espace corporel chez les arthropodes n’est pas le cœlom mais un hémocoel rempli de sang. Au cours du développement embryonnaire de la plupart des arthropodes, des cavités coelomiques vestigiales s’ouvrent dans le mésoderme d’au moins certains somites. Elles sont rapidement oblitérées ou deviennent continues avec l’espace entre les structures mésodermiques et ectodermiques en développement et le vitellus. Cet espace devient l’hémocèle et n’est donc pas tapissé par un péritoine mésodermique. Chez les crustacés, les seuls compartiments coelomiques restants sont les sacs terminaux des organes excréteurs et l’espace autour des gonades.
Système musculaire : Les muscles striés constituent une partie considérable du corps de la plupart des Crustacés. Les muscles sont généralement disposés en groupes antagonistes : les fléchisseurs, qui tirent une partie vers le corps, et les extenseurs, qui l’étendent vers l’extérieur. L’abdomen de l’écrevisse possède de puissants fléchisseurs (illustration 19-5), qui sont utilisés lorsque l’animal nage à reculons, son meilleur moyen de s’échapper. Des muscles puissants situés de part et d’autre de l’estomac contrôlent les mandibules.
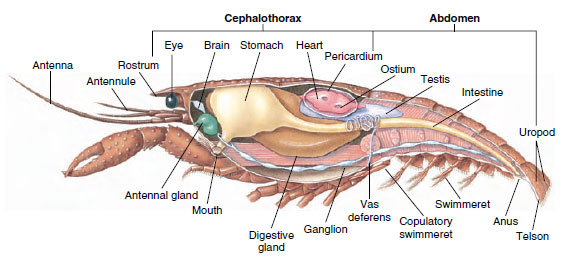
Figure 19-5
Structure interne d’une écrevisse mâle.
Système respiratoire : Les échanges gazeux respiratoires chez les petits crustacés se font sur des zones plus fines de cuticule (par exemple, dans les pattes) ou sur l’ensemble du corps, et les structures spécialisées peuvent être absentes. Les crustacés plus grands ont des branchies, qui sont des projections délicates, ressemblant à des plumes, avec une cuticule très fine. Chez les décapodes, les côtés de la carapace entourent la cavité branchiale, qui est ouverte antérieurement et ventralement (figure 19-6). Les branchies peuvent faire saillie de la paroi pleurale dans la cavité branchiale, de l’articulation des pattes thoraciques avec le corps, ou des coxae thoraciques. Ces deux derniers types sont typiques des écrevisses. L' »écope », une partie du deuxième maxillaire, attire l’eau sur les filaments branchiaux, dans la cavité branchiale à la base des pattes, et hors de la cavité branchiale à la partie antérieure.
Système circulatoire : Les crustacés et autres arthropodes ont un système circulatoire de type « ouvert » ou lacunaire. Cela signifie qu’il n’y a pas de veines ni de séparation du sang et du liquide interstitiel, comme c’est le cas chez les animaux à système fermé. L’hémolymphe (sang) quitte le cœur par les artères, circule dans l’hémocèle et retourne dans les sinus ou espaces veineux, au lieu des veines, avant de revenir dans le cœur. Les annélides ont un système fermé, comme les vertébrés.
Un cœur dorsal est le principal organe de propulsion. Il s’agit d’un sac monochambre de muscle strié. L’hémolymphe pénètre dans le cœur à partir du sinus péricardique environnant par des ostia jumelés, avec des valves qui empêchent le reflux dans le sinus (figure 19-6). À partir du cœur, l’hémolymphe pénètre dans une ou plusieurs artères. Des valves dans les artères empêchent le reflux de l’hémolymphe. Les petites artères se déversent dans les sinus tissulaires, qui à leur tour se déversent souvent dans un grand sinus sternal (figure 19-6).
De là, des canaux sinusaux afférents transportent l’hémolymphe vers les branchies, si elles sont présentes, pour un échange d’oxygène et de dioxyde de carbone. L’hémolymphe retourne ensuite au sinus péricardique par les canaux efférents (figure 19-6).
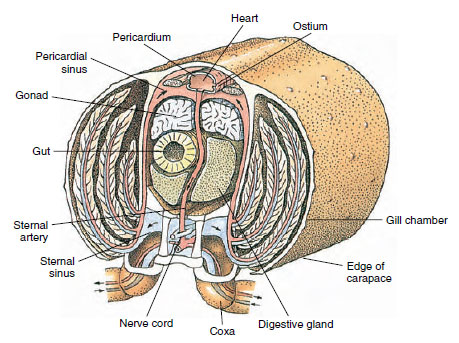
Figure 19-6
Coupe transversale diagrammatique à travers la région cardiaque d’une écrevisse montrant la direction du flux sanguin dans ce
système sanguin « ouvert ». Le cœur pompe le sang vers les tissus de l’organisme par les artères, qui se déversent dans les sinus des tissus. Le
sang de retour entre dans le sinus sternal, puis passe par les branchies pour les échanges gazeux, et enfin retourne au sinus péricardique par des
canaux efférents. Notez l’absence de veines.
L’hémolymphe des arthropodes est en grande partie incolore. Elle comprend des cellules amiboïdes d’au moins deux types. L’hémocyanine, un pigment respiratoire contenant du cuivre, ou l’hémoglobine, un pigment contenant du fer, peuvent être portées en solution. L’hémolymphe a la propriété de coaguler, ce qui empêche sa perte lors de blessures mineures. Certaines cellules amiboïdes libèrent un coagulant de type thrombine qui précipite la coagulation.
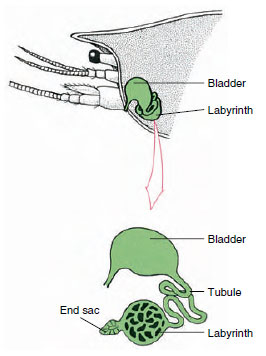
Figure 19-7
Schéma de la glande antennaire (glande verte) des
écrevisses. (En position naturelle, l’organe est beaucoup
plié.) Certains crustacés n’ont pas de labyrinthe,
et le tubule excréteur (canal néphridien) est
un tube très enroulé.
Système excréteur : Les organes excréteurs des crustacés adultes sont une paire de structures tubulaires situées dans la partie ventrale de la tête, en avant de l’œsophage (figure 19-5). On les appelle glandes antennaires ou glandes maxillaires, selon qu’elles s’ouvrent à la base des antennes ou des seconds maxillaires. Quelques crustacés adultes possèdent les deux. Les organes excréteurs des décapodes sont des glandes antennaires, également appelées glandes vertes dans ce groupe. Les crustacés n’ont pas de tubules de Malpighi, les organes excréteurs des araignées et des insectes.
Le sac terminal de la glande antennaire, qui est dérivé d’un compartiment coelomique embryonnaire, se compose d’une petite vésicule (saccule) et d’une masse spongieuse appelée labyrinthe. Le labyrinthe est relié par un tubule excréteur à une vessie dorsale, qui s’ouvre vers l’extérieur par un pore sur la surface ventrale du segment basal de l’antenne (figure 19-7). La pression hydrostatique à l’intérieur de l’hémocèle fournit la force nécessaire à la filtration du fluide dans le sac terminal. Lorsque le filtrat traverse le tubule excréteur et la vessie, il est modifié par la résorption des sels, des acides aminés, du glucose et d’une partie de l’eau et est finalement excrété sous forme d’urine.
L’excrétion des déchets azotés (principalement l’ammoniac) se fait par diffusion à travers de fines zones de cuticule, notamment les branchies, et les organes dits excréteurs fonctionnent principalement pour réguler la composition ionique et osmotique des fluides corporels. Les crustacés d’eau douce, comme les écrevisses, sont constamment menacés de surdilution de leur sang par l’eau, qui diffuse à travers les branchies et autres surfaces perméables à l’eau. Les glandes vertes, en formant une urine diluée et peu salée, agissent comme un dispositif efficace de « contrôle des inondations ». Une partie du Na+ et du Cl- est perdue dans l’urine, mais cette perte est compensée par l’absorption active du sel dissous par les branchies. Chez les crustacés marins, comme les homards et les crabes, le rein a pour fonction d’ajuster la composition en sel de l’hémolymphe par une modification sélective de la teneur en sel de l’urine tubulaire. Dans ces formes, l’urine reste isosmotique au sang.
Systèmes nerveux et sensoriels : Les systèmes nerveux des crustacés et des annélides ont beaucoup en commun, bien que ceux des crustacés présentent plus de fusion de ganglions (figure 19-5). Le cerveau est constitué d’une paire de ganglions supra-œsophagiens qui alimentent en nerfs les yeux et deux paires d’antennes. Il est relié par des connecteurs au ganglion sous-œsophagien, une fusion d’au moins cinq paires de ganglions qui alimentent les nerfs de la bouche, des appendices, de l’œsophage et des glandes antennaires. Le double cordon nerveux ventral comporte une paire de ganglions pour chaque somite et des nerfs desservant les appendices, les muscles et d’autres parties.
En plus de ce système central, il peut y avoir un système nerveux sympathique associé au tube digestif.
Les crustacés ont des organes des sens mieux développés que les annélides. Les plus grands organes des sens des écrevisses sont les yeux et les statocystes. Les poils tactiles, de délicates projections de cuticule, sont largement répartis sur le corps et sont particulièrement abondants sur les chélas, les pièces buccales et le telson. Les sens chimiques du goût et de l’odorat se trouvent dans les poils des antennes, des pièces buccales et d’autres endroits.
Un statocyste en forme de sac, s’ouvrant à la surface par un pore dorsal, se trouve sur le segment basal de chaque première antenne des écrevisses. Le statocyste contient une crête qui porte des poils sensoriels formés à partir du revêtement chitineux et des grains de sable qui servent de statolithes. Chaque fois que l’animal change de position, les changements correspondants dans la position des grains sur les poils sensoriels sont relayés comme stimuli au cerveau, et l’animal peut s’ajuster en conséquence. Chaque mue (ecdysis) de la cuticule entraîne la perte du revêtement cuticulaire du statocyste et avec lui les grains de sable. De nouveaux grains sont ramassés par le pore dorsal après l’ecdysis.
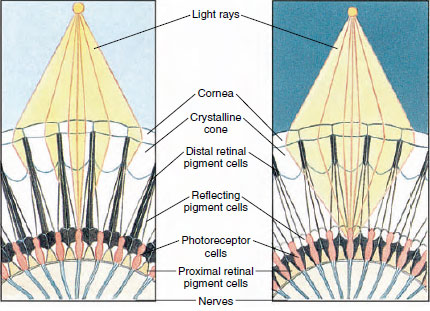
Figure 19-8
Portion d’œil composé d’un arthropode montrant la migration du pigment
dans les ommatidies pour la vision diurne et nocturne. Cinq ommatidies sont représentées dans
chaque diagramme. Le jour, chaque ommatidium est entouré d’un collier
pigmentaire sombre, de sorte que chaque ommatidium n’est stimulé que par les rayons lumineux
qui entrent dans sa propre cornée (vision en mosaïque) ; la nuit, le pigment forme des colliers
incomplets et les rayons lumineux peuvent se propager aux ommatidies
concernées (image continue, ou superposition).
Les yeux de nombreux crustacés sont composés, constitués de nombreuses unités de photorécepteurs appelées ommatidies (figure 19-8). La surface arrondie de chaque œil est recouverte d’une zone transparente de cuticule, la cornée, qui est divisée en de nombreux petits carrés ou hexagones appelés facettes. Ces facettes sont les extrémités extérieures des ommatidies. Chaque ommatidie se comporte comme un petit œil et contient plusieurs types de cellules disposées en colonne (figure 19-8). Les cellules pigmentaires noires se trouvent entre les ommatidies adjacentes.
Le mouvement du pigment dans un œil composé d’arthropode lui permet de s’adapter à différentes quantités de lumière. Il y a trois ensembles de cellules pigmentaires dans chaque ommatidium : rétinal distal, rétinal proximal, et réfléchissant ; ceux-ci sont disposés de telle sorte qu’ils peuvent former un collier ou un manchon plus ou moins complet autour de chaque ommatidium. En cas de forte lumière ou d’adaptation au jour, le pigment rétinien distal se déplace vers l’intérieur et rencontre le pigment rétinien proximal qui se déplace vers l’extérieur, de sorte qu’un manchon pigmentaire complet se forme autour de l’ommatidien (figure 19-8). Dans ces conditions, seuls les rayons qui frappent directement la cornée atteignent les cellules photoréceptrices (rétiniennes), car chaque ommatidium est protégé des autres. Ainsi, chaque ommatidium ne verra qu’une zone limitée du champ de vision (image en mosaïque ou en apposition). Dans une lumière faible, les pigments distaux et proximaux se séparent de sorte que les rayons lumineux, à l’aide des cellules pigmentaires réfléchissantes, ont une chance de se propager aux ommatidies adjacentes et de former une image continue, ou superposition. Ce deuxième type de vision est moins précis mais tire le maximum d’avantages de la quantité limitée de lumière reçue.
Reproduction, cycles de vie et fonction endocrinienne
La plupart des crustacés ont des sexes séparés, et il existe diverses spécialisations pour la copulation parmi les différents groupes. Les balanes sont monoïques mais pratiquent généralement la fécondation croisée. Chez certains ostracodes, les mâles sont rares, et la reproduction est généralement parthénogénétique. La plupart des crustacés couvent leurs œufs d’une manière ou d’une autre : les branchiopodes et les balanes ont des chambres de couvaison spéciales, les copépodes ont des sacs à couvain fixés sur les côtés de l’abdomen (voir figure 19-19), et de nombreux malacostracés portent des œufs et des petits fixés à leurs appendices abdominaux.
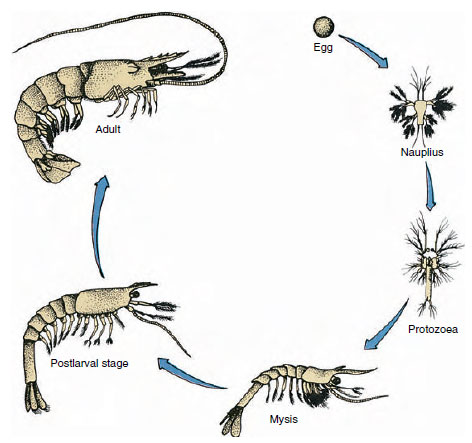
Figure 19-9
Cycle de vie d’une crevette du Golfe Penaeus. Les pénéides fraient à des profondeurs de 40 à 90 m. Les jeunes formes larvaires sont
planktoniques et se déplacent vers les eaux côtières de plus faible salinité pour se développer en tant que juvéniles. Les crevettes plus âgées retournent dans des eaux plus profondes
au large.
Les écrevisses ont un développement direct : il n’y a pas de forme larvaire. Un minuscule juvénile ayant la même forme que l’adulte et un ensemble complet d’appendices et de somites sort de l’œuf. Cependant, le développement est indirect chez la majorité des crustacés, et une larve assez différente de l’adulte par sa structure et son apparence éclot de l’œuf. Le passage de la larve à l’adulte est la métamorphose. La larve primitive et la plus répandue chez les Crustacés est le nauplius (illustrations 19-9 et 19-23). Les nauplies ne portent que trois paires d’appendices : les premières antennes uniramiques, les antennes biramiques et les mandibules biramiques. Tous fonctionnent comme des appendices natatoires à ce stade.
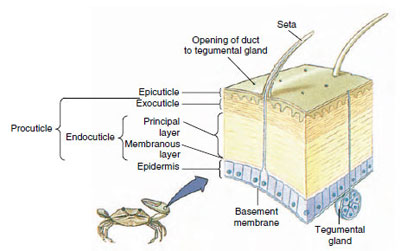
Figure 19-10
Structure de la cuticule des crustacés.
Le développement subséquent peut impliquer un changement graduel vers la forme corporelle adulte, et les appendices et les somites sont ajoutés par une série de mues, ou l’assomption de la forme adulte peut impliquer des changements plus abrupts. Par exemple, la métamorphose d’une balane passe d’un nauplius nageant librement à une larve dotée d’une carapace bivalve appelée cypride et enfin à un adulte sessile doté de plaques calcaires.
Ecdysis : L’ecdysis (ek’duh-sis) (Gr. ekdyein, se dépouiller), ou mue, est nécessaire pour que le corps augmente en taille car l’exosquelette est non vivant et ne grandit pas en même temps que l’animal. Une grande partie du fonctionnement d’un crustacé, y compris sa reproduction, son comportement et de nombreux processus métaboliques, est directement affectée par la physiologie du cycle de mue.
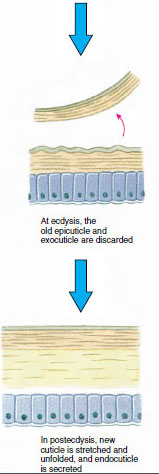
La cuticule, qui est sécrétée par l’épiderme sous-jacent, comporte plusieurs couches (figure 19-10). La plus externe est l’épicuticule, une très fine couche de protéines imprégnées de lipides. L’essentiel de la cuticule est constitué de plusieurs couches de procuticules : (1) l’exocuticule, qui se trouve juste sous l’épicuticule et qui contient des protéines, des sels de calcium et de la chitine ; (2) l’endocuticule, qui est elle-même composée (3) d’une couche principale, qui contient plus de chitine et moins de protéines et qui est fortement calcifiée, et (4) d’une couche membranaire non calcifiée, une couche relativement mince de chitine et de protéines.
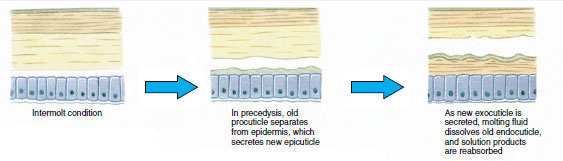
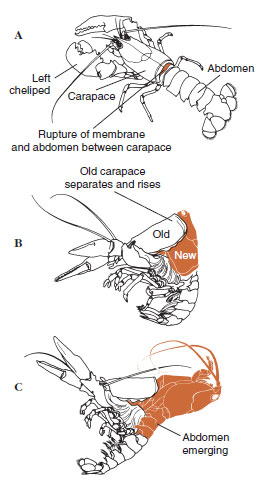
Quelque temps avant l’ecdysis proprement dite, les cellules épidermiques s’agrandissent considérablement. Elles se séparent de la couche membranaire, sécrètent un nouvel épicuticule et commencent à sécréter un nouvel exocuticule (figure 19-11). Des enzymes sont libérées dans la zone située au-dessus de la nouvelle épicuticule. Ces enzymes commencent à dissoudre l’ancienne endocuticule, et les produits solubles sont résorbés et stockés dans le corps du crustacé. Certains sels de calcium sont stockés sous forme de gastrolithes (accrétions minérales) dans les parois de l’estomac. Enfin, il ne reste plus que l’exocuticule et l’épicuticule de l’ancienne cuticule, recouverts par une nouvelle épicuticule et une nouvelle exocuticule. L’animal avale de l’eau, qu’il absorbe par son intestin, et son volume sanguin augmente fortement. La pression interne fait éclater la cuticule et l’animal s’extrait de son ancien exosquelette (illustration 19-12). Suivent alors un étirement de la nouvelle cuticule encore molle, le dépôt de la nouvelle endocuticule, la redéposition des sels inorganiques récupérés et des autres constituants, et le durcissement de la nouvelle cuticule. Pendant la période de mue, l’animal est sans défense et reste caché.
 |
|
 |
 |
| Figure 19-12 Séquence de la mue chez un homard, Homarus americanus. A, la membrane entre la carapace et l’abdomen se rompt, et la carapace commence à s’élever lentement. Cette étape peut prendre jusqu’à 2 heures. B et C, la tête, le thorax et enfin l’abdomen se retirent. Ce processus ne prend généralement pas plus de 15 minutes. Immédiatement après l’ecdysis, les chélipèdes sont desséchés et le corps est très mou. Le homard continue à absorber rapidement de l’eau, de sorte qu’en 12 heures, le corps augmente d’environ 20 % en longueur et 50 % en poids. L’eau des tissus sera remplacée par des protéines au cours des semaines suivantes. |
Figure 19-11 Sécrétion et résorption des cuticules lors de l’ecdysis. |
Lorsqu’un crustacé est jeune, l’ecdysis doit se produire fréquemment pour permettre la croissance, et le cycle de mue est relativement court. Lorsque l’animal approche de la maturité, les périodes d’intermolt deviennent progressivement plus longues, et chez certaines espèces, la mue cesse complètement. Pendant les périodes d’intermolte, une augmentation de la masse tissulaire se produit car les tissus vivants remplacent l’eau.
Contrôle hormonal du cycle d’ecdysis
Bien que l’ecdysis soit contrôlée par les hormones, le cycle est souvent initié par un stimulus environnemental perçu par le système nerveux central. Ces stimuli peuvent inclure la température, la longueur du jour et l’humidité (dans le cas des crabes terrestres). Le signal du système nerveux central diminue la production d’une hormone inhibitrice de la mue par l’organe X. L’organe X est un groupe de cellules neurosécrétoires situées dans la medulla terminalis du cerveau. Chez les écrevisses et autres décapodes, la medulla terminalis se trouve dans le pédoncule oculaire. L’hormone est transportée dans les axones de l’organe X jusqu’à la glande sinus (qui elle-même n’a probablement pas de fonction glandulaire), également dans le pédoncule oculaire, où elle est libérée dans l’hémolymphe.
Une baisse du niveau de l’hormone inhibitrice de la mue favorise la libération d’une hormone de la mue par les organes Y. Les Y-organs se trouvent sous l’épiderme près des muscles adducteurs des mandibules, et ils sont homol-ogus aux glandes prothoraciques des insectes, qui produisent l’hormone ecdysone. L’action de l’hormone de mue est d’initier les processus menant à l’ecdysis (proecdysis). Une fois initié, le cycle se déroule automatiquement sans autre action d’hormones provenant des organes X ou Y.
Autres fonctions endocriniennes : Non seulement l’ablation des pédoncules oculaires accélère la mue, mais on a également découvert il y a plus de 100 ans que les crustacés dont les pédoncules oculaires ont été enlevés ne peuvent plus ajuster la coloration de leur corps aux conditions de fond. Il y a plus de 50 ans, on a découvert que ce défaut n’était pas dû à une perte de vision mais à une perte d’hormones dans les pédoncules oculaires. La couleur du corps des crustacés résulte en grande partie des pigments présents dans des cellules ramifiées spéciales (chromatophores) de l’épiderme.
La concentration des granules de pigment au centre des cellules provoque un effet d’éclaircissement, et la dispersion du pigment dans l’ensemble des cellules provoque un effet d’assombrissement. Le comportement pigmentaire est contrôlé par des hormones provenant des cellules neurosécrétoires du pédoncule oculaire, tout comme la migration du pigment rétinien pour l’adaptation à la lumière et à l’obscurité dans les yeux (figure 19-8).
La libération de matériel neurosécrétoire des organes péricardiques dans la paroi du péricarde provoque une augmentation de la fréquence et de l’amplitude des battements cardiaques.
Des glandes androgènes, découvertes pour la première fois chez un amphipode (Orchestia, un sauteur de plage commun), sont présentes chez les malacostracés mâles. Contrairement à la plupart des autres organes endocriniens des crustacés, ce ne sont pas des organes neurosécrétoires. Leur sécrétion stimule l’expression des caractéristiques sexuelles mâles. Les jeunes malacostracés possèdent des glandes androgènes rudimentaires, mais chez les femelles, ces glandes ne se développent pas. Si elles sont implantées artificiellement chez une femelle, ses ovaires se transforment en testicules et commencent à produire des spermatozoïdes, et ses appendices commencent à prendre des caractéristiques masculines à la mue suivante. Chez les isopodes, les glandes androgènes se trouvent dans les testicules ; chez tous les autres malacostracés, elles se trouvent entre les muscles des coxopodes des dernières pattes thoraciques et sont partiellement fixées près des extrémités des vasa deferentia. Bien que les femelles ne possèdent pas d’organes similaires aux glandes androgènes, leurs ovaires produisent une ou deux hormones qui influencent les caractères sexuels secondaires.
Les hormones qui influencent d’autres processus corporels chez les Crustacés peuvent être présentes, et des preuves suggèrent qu’une substance neurosécrétoire produite dans le pédoncule oculaire régule le niveau de sucre dans le sang.
Habitudes alimentaires
Les habitudes alimentaires et les adaptations à l’alimentation varient beaucoup chez les crustacés. De nombreuses formes peuvent passer d’un type d’alimentation à un autre selon l’environnement et la disponibilité de la nourriture, mais toutes utilisent le même ensemble fondamental de pièces buccales. Les mandibules et les maxillaires servent à ingérer la nourriture ; les maxillipèdes tiennent et écrasent la nourriture. Chez les prédateurs, les pattes mobiles, en particulier les chélipèdes, servent à capturer la nourriture.
De nombreux crustacés, grands et petits, sont prédateurs, et certains ont des adaptations intéressantes pour tuer leurs proies. Une forme semblable à la crevette, Lygiosquilla, possède sur l’une de ses pattes de marche un doigt spécialisé qui peut être tiré dans une rainure et relâché soudainement pour percer une proie qui passe. Les crevettes pistolet (Alpheus spp.) ont une chéla énormément élargie qui peut être armée comme le marteau d’un pistolet et claquée avec une force qui étourdit leur proie.
La nourriture des suspensivores va du plancton et des détritus aux bactéries. Les prédateurs consomment des larves, des vers, des crustacés, des escargots et des poissons. Les charognards mangent des matières animales et végétales mortes. Les suspensivores, comme les crevettes fébriles, les puces d’eau et les balanes, utilisent leurs pattes, qui portent une épaisse frange de soies, pour créer des courants d’eau qui balaient les particules de nourriture à travers les soies. Les crevettes de boue (Upogebia spp.) utilisent de longues soies sur leurs deux premières paires d’appendices thoraciques pour filtrer les matières alimentaires de l’eau circulant dans leur terrier par les mouvements de leurs nageoires.
Les écrevisses ont un estomac en deux parties (figure 19-13). La première partie contient un moulin gastrique dans lequel les aliments, déjà déchiquetés par les mandibules, peuvent être encore broyés par trois dents calcaires en particules suffisamment fines pour passer à travers un filtre sétosique dans la seconde partie ; les particules alimentaires passent ensuite dans l’intestin pour une digestion chimique.
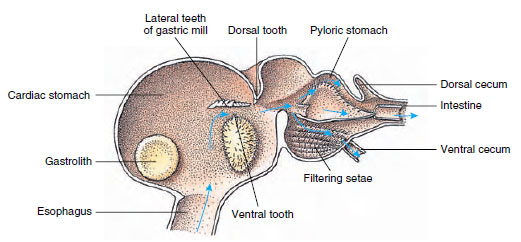
Figure 19-13
L’estomac de Malacostracan montrant le « moulin » gastrique et les directions des mouvements alimentaires. Le moulin possède des crêtes chitineuses, ou
dents, pour la mastication, et des soies pour filtrer les aliments avant leur passage dans l’estomac pylorique.
.
Laisser un commentaire