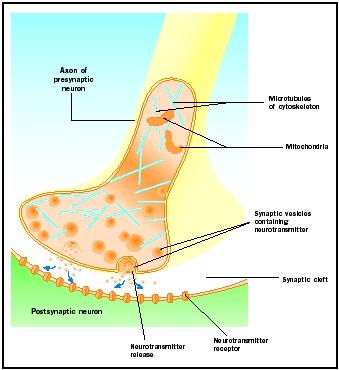
Neurotransmitter Release
Synaptic knobs saapuvat toimintapotentiaalit laukaisevat välittäjäaineen vapautumisen synaptiseen rakoon. Molekyylimekanismia ei täysin ymmärretä. Toimintapotentiaalin saapumisen ja välittäjäaineen vapautumisen välillä on 1-2 millisekunnin ”synaptinen viive”. Toimintapotentiaalit avaavat kalsiumkanavat synaptisen nystyrän kalvossa, mikä aiheuttaa kalsiumionien liikkeen sisäänpäin. Kalsiumionit käynnistävät välittäjäaineen vapautumisen synaptisista rakkuloista synaptiseen rakoon. Synaptiset vesikkelit sulautuvat presynaptiseen kalvoon tämän eksosytoosiprosessin aikana. Vanhojen vesikkelien kalvoista tulee osa presynaptista kalvoa ja uudet vesikkelit irtoavat viereisestä kalvoalueesta. Nämä uudet vesikkelit täytetään myöhemmin uudelleen uusilla syntetisoiduilla tai ”kierrätetyillä” välittäjäaineilla .
Vapautuneet välittäjäaineet diffundoituvat kapean synaptisen raon läpi. Postsynaptisella kalvolla neurotransmitterimolekyylit sitoutuvat kalvoon sitoutuneisiin reseptorimolekyyleihin, joilla on kyseiselle neurotransmitterille spesifiset tunnistuskohdat. Neurotransmitterin sitoutuminen reseptoriin laukaisee kyseiselle reseptorille ominaisen postsynaptisen vasteen. Nämä vasteet voivat olla reseptorin ominaisuuksista riippuen joko kiihdyttäviä tai estäviä. Jos reseptorin stimulaatio johtaa siihen, että postsynaptinen kalvo muuttuu sähköisesti positiivisemmaksi (depolarisoituu), kyseessä on eksitatorinen postsynaptinen potentiaali (EPSP). Jos se muuttuu negatiivisemmaksi (hyperpolarisoituu), kyseessä on inhibitorinen postsynaptinen potentiaali (IPSP). Heräte ja inhibitio riippuvat reseptorin eikä välittäjäaineen ominaisuuksista. Natrium- tai kalsiumkanaviin kytkeytyneet reseptorit ovat eksitatorisia ja saavat aikaan postsynaptisen kalvon depolarisaation, kun taas kloridi- tai kaliumkanaviin kytkeytyneet reseptorit ovat inhibitorisia ja saavat aikaan postsynaptisen kalvon hyperpolarisaation. Tällaisia ionikanaviin kytkettyjä reseptoreita kutsutaan ionotrooppisiksi reseptoreiksi.
Muut reseptorit ovat kytkeytyneet ”second-messenger”-järjestelmiin, jotka käynnistävät sarjan biokemiallisia reaktioita postsynaptisessa solussa. Nämä ovat metabotrooppisia reseptoreita . Metabotrooppiset reseptorit voivat tuottaa monia erilaisia
postsynaptiset tapahtumat. Nämä vaihtelevat vierekkäisten ionikanavien suorasta aktivoitumisesta reseptorien herkkyyden muuttumiseen, tiettyjen lähetti-ribonukleiinihappojen (RNA) transkriptioon tai jopa tiettyjen geenien aktivoitumiseen . Kemialliset synapsit ovat osa hyvin mukautuvaa ja joustavaa viestintäjärjestelmää. Ne eivät ole staattisia anatomisia rakenteita, joilla on kiinteitä ominaisuuksia, vaan dynaamisia rakenteita, jotka pystyvät muuttamaan molekyyliominaisuuksiaan olosuhteiden muuttuessa.
Neurotransmittereita on kirjaimellisesti satoja. Jotkut ovat melko yksinkertaisia yhdisteitä, kuten asetyylikoliini, serotoniini, katekoliamiinit (dopamiini, noradrenaliini ja adrenaliini) ja monet aminohapot . Monet ovat monimutkaisempia ja kuuluvat laajaan joukkoon neuropeptidilähettimiä. Kun neurotransmitterit on vapautettu synaptiseen rakoon, ne pysyvät aktiivisina, kunnes ne joko muuttuvat kemiallisesti tai erityiset kuljetinjärjestelmät ottavat ne takaisin synaptiseen nuppuun ja kierrättävät ne. Kolinergisissä synapseissa asetyylikoliiniesteraasia on synaptisessa raossa. Tämä entsyymi pilkkoo välittäjäaineen asetaatiksi ja koliiniksi, joista kumpikaan ei ole aktiivinen. Serotoniini ja adrenaliini sen sijaan otetaan presynaptiseen terminaaliin ja kierrätetään.
Vastaa