Subphylum Crustacea
General Nature of a Crustacea
Crustaceans differoi muista niveljalkaisista monin eri tavoin, mutta ainoa todella erottuva piirre on se, että äyriäiset ovat ainoat niveljalkaiset, joilla on kaksi paria antenneja. Kahden antenniparin ja alaleukaparin lisäksi äyriäisillä on päässään kaksi paria leukoja, ja sen jälkeen jokaisessa ruumiinosassa eli somitissa on pari lisäkkeitä. Joillakin äyriäisillä ei kaikilla somiteilla ole lisäkkeitä. Kaikki lisäkkeet, ehkä ensimmäisiä antenneja lukuun ottamatta, ovat alkuaan kaksihaaraisia (kaksi päähaaraa), ja ainakin joissakin nykyisten aikuisten lisäkkeissä on tämä tila. Hengitykseen erikoistuneet elimet, jos niitä on, toimivat kiduksina.
Useimmilla äyriäisillä on 16-20 somitiittia, mutta joillakin muodoilla on 60 somitiittia tai enemmän. Suurempi määrä somiitteja on alkukantainen piirre. Johdetumpi tila on se, että segmenttejä on vähemmän ja tagmatisaatio on lisääntynyt. Tärkeimmät tagmat ovat pää, rintakehä ja vatsa, mutta nämä eivät ole homologisia koko luokassa (tai edes joidenkin alaluokkien sisällä), koska somiittien sulautumisasteet vaihtelevat, kuten esimerkiksi cephalothoraxissa.
Kuva 19-1
Malacostracan arkkityyppinen kuvio. Kaksi leukaluuta ja kolme
leukaluuta on erotettu kaavamaisesti havainnollistamaan
yleissuunnitelmaa.
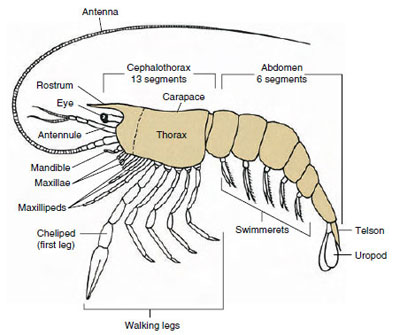
Ylivoimaisesti suurin äyriäisryhmä on luokka Malacostraca, johon kuuluvat hummerit, ravut, ravut, katkaravut, rantahyppääjät, emakkohyppääjät, emakkohyppääjät ja monet muut. Näissä esiintyy yllättävän vakioitu ruumiinsegmenttien ja tagmata-järjestys, jota pidetään luokan esi-isäkaaviona (kuva 19-1). Tässä tyypillisessä ruumiinrakenteessa pää koostuu viidestä (alkiomaisesti kuudesta) sulautuneesta somiitista, rintakehä kahdeksasta somiitista ja vatsa kuudesta somiitista (muutamilla lajeilla seitsemästä). Etupäässä on segmentoimaton rostrum ja takapäässä segmentoimaton telson, joka viimeisen vatsasomiitin ja sen uropodien kanssa muodostaa monissa muodoissa pyrstöviuhkan.
Monilla äyriäisillä pään selkäkudos voi ulottua taaksepäin ja eläimen sivujen ympärille peittämään tai sulautumaan yhteen joidenkin tai kaikkien rinta- ja vatsasomiittien kanssa. Tätä suojusta kutsutaan karapaxiksi. Joissakin ryhmissä carapace muodostaa simpukkamaiset läpät, jotka peittävät suurimman osan tai koko ruumiin. Kymmenjalkaisilla (mukaan lukien hummerit, katkaravut, ravut ja muut) selkäkilpi peittää koko kefalothoraxin, mutta ei vatsaa.
Muoto ja toiminta
Kokonsa ja helpon saatavuutensa vuoksi suuria äyriäisiä, kuten rapuja, on tutkittu enemmän kuin muita ryhmiä. Niitä tutkitaan yleisesti myös johdantolaboratoriokursseilla. Siksi monet seuraavassa esitetyistä huomioista koskevat nimenomaan rapuja ja niiden sukulaisia.
Ulkoiset piirteet
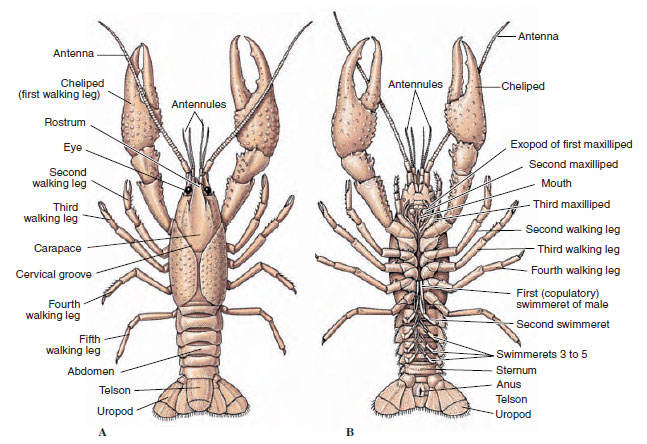
Katkarapujen vartaloa peittää erittyvä kutikula, joka koostuu kitiinistä, proteiinista ja kalkkipitoisesta aineesta. Suurempien äyriäisten kovemmissa, painavissa levyissä on erityisen paljon kalkkipitoisia kerrostumia. Kova suojakuori on pehmeä ja ohut somiittien välisissä nivelissä, mikä mahdollistaa liikkumisen joustavuuden. Mahdollinen selkäkilpi peittää suuren osan tai koko kefalothoraxin; kymmenjalkaisilla, kuten ravuilla, kaikki pään ja rintakehän segmentit ovat selkäpuolelta selkäkilven ympäröimiä. Jokaista somitea, joka ei ole selkäkilven ympäröimä, peittää dorsaalinen cutikulaarilevy eli tergum (kuva 19-2A), ja segmenttiläiskien välissä on ventraalinen poikkipalkki eli sternum (kuva 19-2B). Vatsa päättyy telsoniin, jota ei pidetä somiittina ja jossa on peräaukko. (Telson saattaa olla homologinen annelidien pygidiumin kanssa.)
Kuva 19-2
Rapujen ulkoinen rakenne. A, Selkäkuva. B, Ventraalinen näkymä.
Kuva 19-3
Biramous-äyriäisen
liitännäisen
osat (
ravun
kolmannen maxillipedin
päällysosa)
Gonoporeiden asento vaihtelee sukupuolestaan ja äyriäisryhmästänsä riippuen. Ne voivat olla umpisolmuparin päällä tai sen tyvessä, ruumiin loppupäässä tai ilman jalkoja olevissa somiteissa. Rapuilla vasa deferentian aukot ovat keskisivulla viidennen kävelyjalkaisen parin tyvessä, ja munanjohtimien aukot ovat kolmannen parin tyvessä. Naaraiden siemennesteen aukko sijaitsee yleensä neljännen ja viidennen kävelyjalkaparin välisellä keskiviivalla.
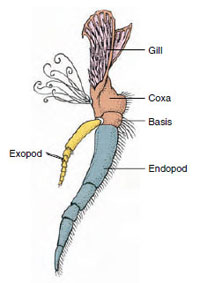
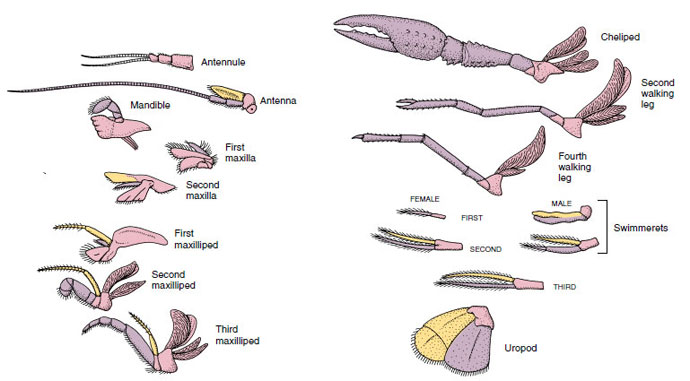
Lisäkkeet: Malacostraca-luokkien (mukaan lukien ravut) ja Remipedia-luokkien jäsenillä on tyypillisesti pari nivellettyä lisäkettä jokaisessa somiteessa (kuva 19-3), vaikka muiden luokkien vatsasomiiteissa ei ole lisäkkeitä. Huomattavaa erikoistumista on havaittavissa johdettujen äyriäisten, kuten rapujen, umpilisäkkeissä. Kaikki ovat kuitenkin variaatioita biramous-perussuunnitelmasta, jota havainnollistaa rapujen lisäkkeet, kuten maxilliped (rintakehän raaja, joka on muunnettu pään lisäkkeeksi) (kuvat 19-3 ja 19-4). Tyviosassa eli protopodissa on lateraalinen exopodi ja mediaalinen endopodi. Protopodi koostuu yhdestä tai kahdesta nivelestä (basis ja coxa), kun taas exopodissa ja endopodissa on kummassakin yhdestä useampaan niveleen. Joistakin lisäkkeistä, kuten rapujen kävelevistä jaloista, on tullut sekundaarisesti yksijalkaisia. Äyriäisten raajoissa esiintyy joskus mediaalisia tai lateraalisia ulokkeita, joita kutsutaan vastaavasti endiiteiksi ja eksitiiteiksi, ja protopodin eksitiittiä kutsutaan epipodiksi. Epipodit ovat usein muuntuneet kiduksiksi. Taulukossa 19-1 on esitetty, miten eri lisäkkeet ovat muuntuneet biramous-kaavasta tiettyihin toimintoihin sopiviksi.
Kuva 19-4
Ravun lisäkkeet, joista käy ilmi, miten ne ovat muuntuneet biramous-perussuunnitelmasta, kuten
swimmeretillä. Protopodi, vaaleanpunainen; endopodi, violetti; eksopodi, keltainen.
Rakenteita, joilla on samanlainen perussuunnitelma ja jotka ovat polveutuneet yhteisestä muodosta, sanotaan homologisiksi riippumatta siitä, onko niillä sama tehtävä vai ei. Koska erikoistuneet kävelevät jalat, suulakit, leukalihakset ja uimarit ovat kaikki kehittyneet yhteisestä biramotyypistä, mutta ne ovat muuntuneet suorittamaan eri toimintoja, ne ovat kaikki homologisia keskenään, mikä tunnetaan nimellä sarjahomologia. Alun perin raajat olivat kaikki hyvin samankaltaisia, mutta rakennemuutosten evoluution aikana osa haaroista on vähentynyt, osa kadonnut, osa muuttunut huomattavasti ja joitakin uusia osia on lisätty. Rapuilla ja niiden liittolaisilla on eläinkunnan kehittynein sarjahomologia, sillä niillä on 17 erilaista, mutta sarjahomologista lisäketyyppiä (Taulukko 19-1).
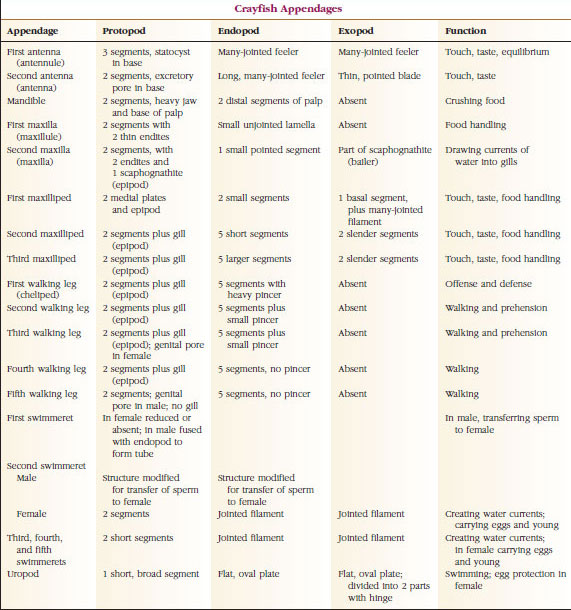
TAULUKKO 19.1
Sisäiset piirteet
Lihas- ja hermojärjestelmissä sekä rintakehän ja vatsan segmentoitumisessa on selvästi havaittavissa annelidien kaltaisilta esi-isiltä periytynyttä metameralismia, mutta muissa järjestelmissä on selviä muutoksia. Useimmissa muutoksissa on kyse osien keskittymisestä tietylle alueelle tai sitten osien, kuten intersepta, vähenemisestä tai täydellisestä häviämisestä.
Hemocoel: Niveljalkaisten tärkein ruumiin tila ei ole kotelo, vaan veren täyttämä hemocoel. Useimpien niveljalkaisten alkionkehityksen aikana ainakin joidenkin somiittien mesodermissä avautuu vestigiaalisia koteloonteloita. Ne häviävät pian tai muuttuvat jatkuviksi kehittyvien mesodermi- ja ektodermirakenteiden ja keltarauhasen välisen tilan kanssa. Tästä tilasta tulee hemokeli, eikä sitä siten vuoraa mesoderminen vatsakalvo. Äyriäisissä ainoat jäljellä olevat coelomiset lokerot ovat erittävien elinten loppupussit ja tila sukurauhasten ympärillä.
Muscular System: Raidalliset lihakset muodostavat huomattavan osan useimpien äyriäisten kehosta. Lihakset on yleensä järjestetty vastakkaisiin ryhmiin: taivuttajiin, jotka vetävät osaa kohti kehoa, ja ojentajiin, jotka ojentavat sitä ulospäin. Rapujen vatsassa on voimakkaat taivuttajat (kuva 19-5), joita käytetään eläimen uidessa taaksepäin, mikä on sen paras pakokeino. Vahvat lihakset vatsan molemmin puolin ohjaavat alaleukoja.
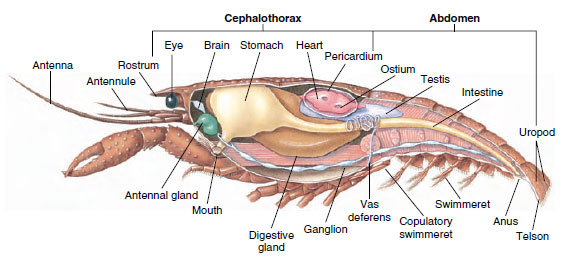
Kuva 19-5
Urosravun sisäinen rakenne.
Hengityselimet: Pienempien äyriäisten hengityskaasujen vaihto tapahtuu ohuemmilla kynsinauhan alueilla (esimerkiksi jaloissa) tai koko kehossa, ja erikoistuneet rakenteet voivat puuttua. Suuremmilla äyriäisillä on kidukset, jotka ovat herkkiä, höyhenenkaltaisia ulokkeita, joissa on hyvin ohut kynsinauha. Kymmenjalkaisilla selkäkilven sivut sulkevat sisäänsä kidusten ontelon, joka on avoin etu- ja vatsapuolelta (kuva 19-6). Kylkiluut voivat työntyä keuhkopussin seinämästä kidusten onteloon, rintakehän jalkojen ja rungon välisestä nivelestä tai rintakehän koakseleista. Kaksi jälkimmäistä tyyppiä ovat tyypillisiä ravuille. Toisen yläleuan osa, ”bailer”, vetää vettä kidussäikeiden yli, jalkojen tyvestä kidusten onteloon ja etupuolelta ulos kidusten ontelosta.
Kiertoelimistö: Äyriäisillä ja muilla niveljalkaisilla on ”avoin” eli lakunaarinen verenkiertojärjestelmä. Tämä tarkoittaa, että veressä ei ole suonia eikä veren ja interstitiaalisen nesteen erottelua, kuten eläimillä, joilla on suljettu järjestelmä. Hemolymfa (veri) poistuu sydämestä valtimoita pitkin, kiertää hemocoelin läpi ja palaa laskimoiden sijasta laskimoiden sinuksiin tai tiloihin, ennen kuin se palaa takaisin sydämeen. Selkärankaisilla on suljettu järjestelmä, kuten selkärankaisillakin.
Selkäydin on tärkein käyttövoimaelin. Se on yksikammioinen pussi, joka koostuu raidallisesta lihaksesta. Hemolymfa kulkeutuu sydämeen ympäröivästä sydänpussinuksesta parittaisten ostioiden kautta, joissa on venttiilit, jotka estävät takaisinvirtauksen sinukseen (kuva 19-6). Sydämestä hemolympha kulkeutuu yhteen tai useampaan valtimoon. Valtimoissa olevat venttiilit estävät hemolymfan takaisinvirtauksen. Pienet valtimot tyhjenevät kudossinuksiin, jotka puolestaan usein purkautuvat suureen rintalastan sinukseen (kuva 19-6).
Sieltä afferentit sinuskanavat kuljettavat hemolymfaa kiduksiin, jos niitä on, hapen ja hiilidioksidin vaihtoa varten. Hemolymfa palaa sitten efferenttejä kanavia pitkin takaisin perikardiaaliseen sinukseen (kuva 19-6).
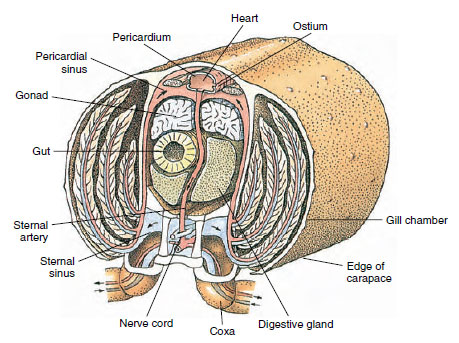
Kuva 19-6
Diagrammattinen poikkileikkaus ravun sydämen alueen läpi, josta käy ilmi veren virtaussuunta tässä ”avoimessa”
verenkierrossa. Sydän pumppaa verta elimistön kudoksiin valtimoiden kautta, jotka tyhjenevät kudossuoniin. Palautuva
veri kulkeutuu rintalastan sivuonteloon, sen jälkeen kidusten läpi kaasujen vaihtoa varten ja lopulta takaisin sydänpussin sivuonteloon efferenttejä
kanavia pitkin. Huomaa laskimoiden puuttuminen.
Niveljalkaisten hemolymfa on suurelta osin väritöntä. Se sisältää ameboidisoluja, joita on ainakin kahta tyyppiä. Hemosyaniinia, kuparia sisältävää hengityspigmenttiä, tai hemoglobiinia, rautaa sisältävää pigmenttiä, voi olla liuoksessa. Hemolymfalla on hyytymisominaisuus, joka estää sen häviämisen pienissä vammoissa. Jotkut ameboidisolut vapauttavat trombiinin kaltaista hyytymisaineita, jotka saostavat hyytymistä.
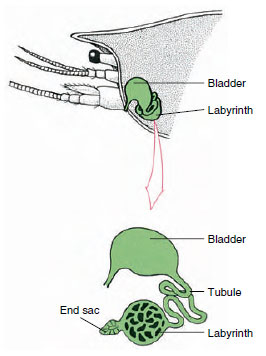
Kuva 19-7
Rapujen antennirauhasen (vihreä rauhanen) järjestelmä. (Luonnollisessa asennossa elin on paljon
taitettu.) Joiltakin äyriäisiltä puuttuu labyrintti,
ja erittävä putki (nephridiaalikanava) on
paljonkierretty putki.
Ekretorinen järjestelmä: Aikuisten äyriäisten erityselimet ovat pari putkimaista rakennetta, jotka sijaitsevat pään ventraalisessa osassa ruokatorven etupuolella (kuva 19-5). Niitä kutsutaan antennirauhasiksi tai leukarauhasiksi riippuen siitä, avautuvatko ne antennien vai toisen leukarauhasen tyvestä. Muutamilla aikuisilla äyriäisillä on molemmat. Kymmenjalkaisten ulostuselimet ovat antennirauhasia, joita kutsutaan tässä ryhmässä myös vihreiksi rauhasiksi. Äyriäisillä ei ole malpighian tubuluksia, jotka ovat hämähäkkien ja hyönteisten erittäviä elimiä.
Alkioperäisestä keliosastosta peräisin olevan antennirauhasen loppupussi koostuu pienestä rakkulasta (saccule) ja sienimäisestä massasta, jota kutsutaan labyrintiksi. Labyrintti yhdistyy erittävän putkilon kautta dorsaaliseen rakkoon, joka avautuu ulospäin antennin tyvisegmentin ventraalipinnalla olevan huokosen kautta (kuva 19-7). Hydrostaattinen paine hemocoelin sisällä antaa voiman nesteen suodattamiselle loppupussiin. Kun suodos kulkee erittävän tubuluksen ja virtsarakon läpi, se muuttuu suolojen, aminohappojen, glukoosin ja jonkin verran veden resorptiolla ja erittyy lopulta virtsana.
Typpipitoisten jätteiden (enimmäkseen ammoniakin) erittyminen tapahtuu diffuusiolla ohuiden kynsinauhojen, erityisesti kidusten, läpi, ja niin kutsuttujen erittävien elinten tehtävänä on pääasiassa säätää elimistön nesteiden ionista ja osmoottista koostumusta. Makean veden äyriäisiä, kuten rapuja, uhkaa jatkuvasti veren liiallista laimentumista vedestä, joka diffundoituu kidusten ja muiden vettä läpäisevien pintojen läpi. Vihreät rauhaset, jotka muodostavat laimeaa, vähäsuolaista virtsaa, toimivat tehokkaana ”tulvanhallintalaitteena”. Virtsan mukana menetetään jonkin verran Na+- ja Cl-arvoja, mutta tämä menetys korvataan aktiivisella liuenneen suolan imeytymisellä kiduksiin. Merieläimissä, kuten hummereissa ja ravuissa, munuaisten tehtävänä on säätää hemolymfan suolakoostumusta muuttamalla valikoivasti tubulaarisen virtsan suolapitoisuutta. Näissä muodoissa virtsa pysyy isosmoottisena vereen nähden.
Hermo- ja aistijärjestelmät: Äyriäisten ja annelidien hermostossa on paljon yhteistä, vaikka äyriäisten hermostossa on enemmän ganglioiden sulautumia (kuva 19-5). Aivoissa on pari supraesofageaalista gangliota, jotka syöttävät hermoja silmille ja kahdelle antenniparille. Se on yhdistetty sidekudoksilla subesofageaaliseen ganglioon, joka on vähintään viiden ganglioparin fuusio, joka syöttää hermoja suuhun, lisäkkeisiin, ruokatorveen ja antennirauhasiin. Kaksinkertaisessa ventraalisessa hermosäikeessä on gangliopari kutakin somitiittia kohti ja hermoja, jotka palvelevat umpisolmukkeita, lihaksia ja muita osia.
Tämän keskusjärjestelmän lisäksi voi olla ruuansulatuskanavaan liittyvä sympaattinen hermosto.
Käpyeläimillä on paremmin kehittyneet aistielimet kuin annelioideilla. Rapujen suurimmat aistielimet ovat silmät ja statokystat. Tuntokarvat ovat laajalti levinneet ympäri kehoa, ja ne ovat hienovaraisia kynsinauhan ulokkeita, joita on erityisen runsaasti kitalakeissa, suulakkeissa ja telsonissa. Kemialliset maku- ja hajuaistit ovat karvoissa muun muassa antenneissa ja suulakkeissa.
Rapujen jokaisen ensimmäisen antennin tyvisegmentissä on pussimainen statokysta, joka avautuu pintaan dorsaalisen huokosen kautta. Statokysta sisältää harjanteen, jossa on kitiinipitoisesta limakalvosta muodostuneita aistikarvoja ja hiekanjyviä, jotka toimivat statoliitteina. Aina kun eläin muuttaa asentoaan, vastaavat muutokset aistikarvojen rakeiden asennossa välittyvät ärsykkeinä aivoihin, ja eläin voi mukautua sen mukaisesti. Jokainen kutikulaarinen sulatus (ekdyysi) johtaa statokystan kutikulaarisen limakalvon ja sen mukana hiekanjyvien katoamiseen. Uudet jyvät poimitaan dorsaalisen huokosen kautta ekdysiksen jälkeen.
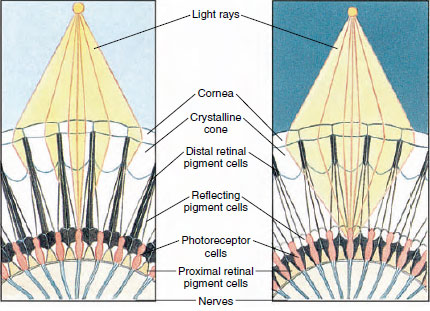
Kuva 19-8
Partikkeli niveljalkaisen yhdyssilmästä, jossa näkyy pigmentin
siirtyminen
ommatidioissa päivä- ja yönäköä varten. Kussakin kuvassa on viisi ommatidiaa. Päivällä kutakin ommatidiaa ympäröi tumma
pigmenttikaulus, joten kutakin ommatidiaa stimuloivat vain valonsäteet
, jotka tulevat sen omaan sarveiskalvoon (mosaiikkinäkö); yöllä pigmentti muodostaa
epätäydelliset kaulukset, ja valonsäteet voivat levitä viereisiin ommatidioihin
(jatkuva eli päällekkäinen kuva).
Monien äyriäisten silmät ovat yhdistelmäsilmiä, jotka koostuvat monista ommatidioiksi kutsutuista valoreseptoriyksiköistä (kuva 19-8). Kunkin silmän pyöristettyä pintaa peittää läpinäkyvä kynsinauhan alue, sarveiskalvo, joka on jaettu moniin pieniin neliöihin tai kuusikulmioihin, joita kutsutaan faseteiksi. Nämä fasetit ovat ommatidioiden uloimpia päitä. Kukin ommatidium käyttäytyy kuin pieni silmä, ja se sisältää useita erilaisia soluja, jotka on järjestetty pylväsmäisesti (kuva 19-8). Vierekkäisten ommatidioiden välissä on mustia pigmenttisoluja.
Niveljalkaisten yhdistelmäsilmän pigmenttisolujen liikkeen ansiosta se pystyy sopeutumaan erilaisiin valomääriin. Jokaisessa ommatidiumissa on kolme pigmenttisoluryhmää: distaalinen verkkokalvo, proksimaalinen verkkokalvo ja heijastussolut; ne on järjestetty siten, että ne voivat muodostaa enemmän tai vähemmän täydellisen kauluksen tai holkin jokaisen ommatidiumin ympärille. Voimakkaassa valossa tai päiväadaptaatiossa distaalinen verkkokalvopigmentti siirtyy sisäänpäin ja kohtaa ulospäin liikkuvan proksimaalisen verkkokalvopigmentin niin, että ommatidiumin ympärille muodostuu täydellinen pigmenttiholkki (kuva 19-8). Tässä tilassa vain sarveiskalvoon suoraan osuvat säteet saavuttavat valoreseptorisolut (retinulaariset solut), sillä kukin ommatidium on suojattu muilta. Näin kukin ommatidium näkee vain rajoitetun alueen näkökentästä (mosaiikki- eli appositio-kuva). Hämärässä valossa distaaliset ja proksimaaliset pigmentit erottuvat toisistaan niin, että valonsäteillä on heijastavien pigmenttisolujen avulla mahdollisuus levitä viereisiin ommatidioihin ja muodostaa jatkuva eli päällekkäinen kuva. Tämä jälkimmäinen näkökyky on epätarkempi, mutta se hyödyntää mahdollisimman hyvin vastaanotetun valon rajallista määrää.
Reproduktio, elinkaari ja hormonitoiminta
Useimmilla äyriäisillä on erilliset sukupuolet, ja eri ryhmissä on erilaisia erikoistumismahdollisuuksia paritteluun. Lahnat ovat yksikotisia, mutta harjoittavat yleensä ristiinhedelmöitymistä. Joillakin ostrakodeilla uroksia on vähän, ja lisääntyminen on yleensä parthenogeneettistä. Useimmat äyriäiset hautovat muniaan jollakin tavalla: haarapääjalkaisilla ja simpukoilla on erityisiä hautomakammioita, kopepodeilla on vatsan sivuille kiinnitettyjä hautomapusseja (ks. kuva 19-19), ja monet malakostrakat kantavat munia ja poikasia vatsan ulokkeisiinsa kiinnitettynä.
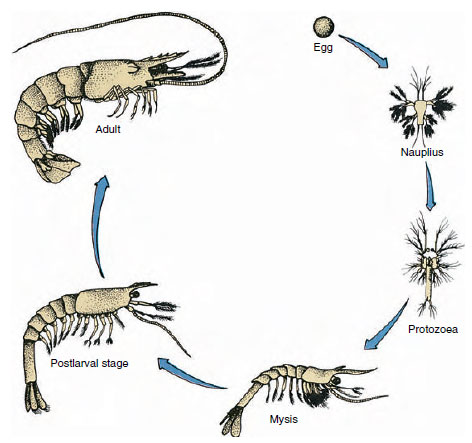
Kuva 19-9
Lahden katkaravun Penaeus elinkaari. Penaeidit kutevat 40-90 m:n syvyydessä. Nuoret toukkamuodot ovat
planktonisia ja siirtyvät rannikolle matalamman suolapitoisuuden omaavaan veteen kehittyäkseen nuoriksi yksilöiksi. Vanhemmat katkaravut palaavat syvempään veteen
rannikolle.
Ravut kehittyvät suoraan: toukkamuotoa ei ole. Munasta kuoriutuu pikkuruinen poikanen, jolla on sama muoto kuin aikuisella ja jolla on täydelliset lisäkkeet ja somitit. Useimmilla äyriäisillä kehitys on kuitenkin epäsuoraa, ja munasta kuoriutuu toukka, joka on rakenteeltaan ja ulkonäöltään täysin erilainen kuin aikuinen. Muutos toukasta lopulta aikuiseksi on metamorfoosi. Äyriäisten alkeellinen ja yleisimmin esiintyvä toukka on nauplius (kuva 19-9 ja 19-23). Nauplioilla on vain kolme paria lisäkkeitä: yksipuoliset ensimmäiset antennit, kaksipuoliset antennit ja kaksipuoliset alaleuat. Kaikki toimivat tässä vaiheessa uivina lisäkkeinä.
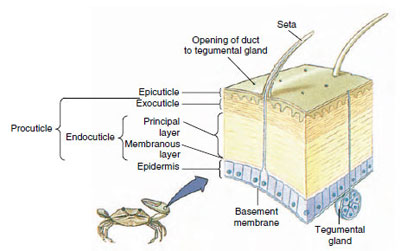
Kuva 19-10
Äyriäisten kynsinauhan rakenne.
Seuraavaan kehitykseen voi sisältyä asteittainen muutos aikuisen ruumiin muotoon, ja lisäkkeet ja somitit lisätään useiden molttien kautta, tai aikuisen muodon omaksumiseen voi liittyä äkillisempiä muutoksia. Esimerkiksi simpukan metamorfoosi etenee vapaasti uivasta naupliuksesta toukaksi, jolla on simpukkapanssari, jota kutsutaan sypridiksi, ja lopulta sessiiliksi aikuiseksi, jolla on kalkkilevyt.
Ecdysis: Ekdyysi (ek´duh-sis) (kreik. ekdyein, riisua) eli moltisoituminen on välttämätöntä kehon koon kasvattamiseksi, koska ulkoluuranko on eloton eikä kasva eläimen kasvaessa. Suuri osa äyriäisen toiminnasta, mukaan lukien sen lisääntyminen, käyttäytyminen ja monet aineenvaihduntaprosessit, on suoraan riippuvainen molting-syklin fysiologiasta.
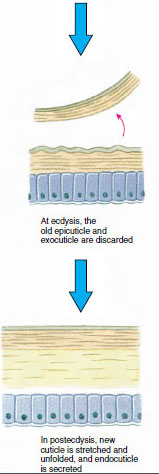
Kutikula, joka erittyy sen alla olevasta epidermiksestä, on monikerroksinen (kuva 19-10). Uloin on epikutikula, hyvin ohut kerros lipidillä kyllästettyä proteiinia. Suurin osa kynsinauhasta on useita kerroksia prokutiikkia: (1) eksokutikula, joka on juuri epikutiklan alapuolella ja sisältää proteiinia, kalsiumsuoloja ja kitiiniä; (2) endokutikula, joka puolestaan koostuu (3) pääkerroksesta, joka sisältää enemmän kitiiniä ja vähemmän proteiinia ja on voimakkaasti kalkkeutunut, ja (4) kalkkeutumattomasta kalvokerroksesta, joka on suhteellisen ohut kitiiniä ja proteiinia sisältävä kerros.
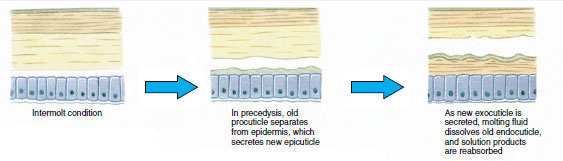
Joitakin aikoja ennen varsinaista ekdyysiä epidermaaliset solut suurentuvat huomattavasti. Ne irtoavat kalvokerroksesta, erittävät uutta epikutiilia ja alkavat erittää uutta eksokutiilia (kuva 19-11). Entsyymejä vapautuu uuden epikutiilin yläpuolelle. Nämä entsyymit alkavat liuottaa vanhaa endokutikkelia, ja liukoiset tuotteet resorboituvat ja varastoituvat äyriäisen kehoon. Osa kalsiumsuoloista varastoituu mahalaukun seinämiin gastroliteiksi (mineraalikertymiksi). Lopulta vanhasta kotelosta jää jäljelle vain eksokutikula ja epikutikula, joiden alla on uusi epikutikula ja uusi eksokutikula. Eläin nielee vettä, jota se imee suoliston kautta, ja sen veren määrä kasvaa huomattavasti. Sisäinen paine saa aikaan sen, että kutikula halkeaa, ja eläin vetää itsensä ulos vanhasta ulkoluurangostaan (kuva 19-12). Tämän jälkeen seuraa vielä pehmeän uuden kitalakihon venyminen, uuden endokutiklan laskeutuminen, pelastettujen epäorgaanisten suolojen ja muiden ainesosien uudelleen laskeutuminen ja uuden kitalakihon kovettuminen. Moltingin aikana eläin on puolustuskyvytön ja pysyy piilossa.
 |
|
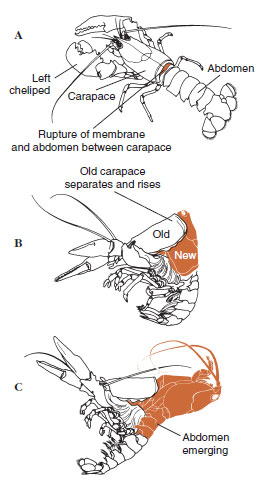 |
 |
| Kuvio 19-12 Sulamisjärjestys hummerissa, Homarus americanus. A, Karapaxin ja vatsan välinen kalvo repeää, ja karapax alkaa hitaasti kohota. Tämä vaihe voi kestää jopa 2 tuntia. B ja C, pää, rintakehä ja lopulta vatsa vetäytyvät. Tämä prosessi kestää yleensä enintään 15 minuuttia. Välittömästi ekdysiksen jälkeen jalkaterät ovat kuivuneet ja ruumis on hyvin pehmeä. Hummeri jatkaa nopeaa veden imeytymistä niin, että 12 tunnin kuluessa ruumiin pituus kasvaa noin 20 % ja paino 50 %. Kudosvesi korvataan proteiinilla seuraavien viikkojen aikana. |
Kuva 19-11 Kudoksen erittyminen ja resorptio ekdysiksessä. |
Kun äyriäinen on nuori, ekdyssiä on tapahduttava usein, jotta kasvu olisi mahdollista, ja molttijakso on suhteellisen lyhyt. Kun eläin lähestyy sukukypsyyttä, intermolttijaksot pitenevät asteittain, ja joillakin lajeilla moltti loppuu kokonaan. Intermolttijaksojen aikana kudosmassa kasvaa, kun elävä kudos korvaa vettä.
Ekodyysisyklin hormonaalinen ohjaus
Vaikka ekdyysiä ohjataan hormonaalisesti, sykli käynnistyy usein keskushermoston havaitsemasta ympäristöärsykkeestä. Tällaisia ärsykkeitä voivat olla lämpötila, päivän pituus ja kosteus (maakrapujen tapauksessa). Keskushermostosta tuleva signaali vähentää homehtumista estävän hormonin tuotantoa X-elimessä. X-elin on ryhmä hermosolujen erittäviä soluja aivojen medulla terminaliksessa. Ravuilla ja muilla kymmenjalkaisilla medulla terminalis sijaitsee silmävarressa. Hormoni kulkeutuu X-elimen aksoneissa sinusrauhaseen (joka itsessään ei luultavasti ole rauhanen), joka sijaitsee niin ikään silmänvarressa, jossa se vapautuu hemolymfaan.
Sulautumishormonin tason lasku edistää sulautumishormonin vapautumista Y-elimistä. Y-orgaanit ovat epidermiksen alapuolella lähellä mandibuloiden adductor-lihaksia, ja ne ovat homol-ogus hyönteisten prothorakaalisten rauhasten kanssa, jotka tuottavat ekdysonihormonia. Molttihormonin tehtävänä on käynnistää ekdyysiin (proekdyysiin) johtavat prosessit. Kun sykli on käynnistynyt, se etenee automaattisesti ilman X- tai Y-elinten hormonien vaikutusta.
Muut endokriiniset toiminnot: Sen lisäksi, että silmänvarsien poistaminen nopeuttaa moltingia, havaittiin yli 100 vuotta sitten, että äyriäiset, joiden silmänvarret on poistettu, eivät enää pysty mukauttamaan ruumiin väritystä taustaolosuhteiden mukaan. Yli 50 vuotta sitten havaittiin, että vika ei johdu näkökyvyn heikkenemisestä vaan hormonien häviämisestä silmänvarresta. Äyriäisten ruumiinväri johtuu suurelta osin epidermiksen erityisten haaroittuneiden solujen (kromatofoorien) pigmenteistä.
Pigmenttirakeiden keskittyminen solujen keskelle aiheuttaa vaalentavan vaikutuksen, ja pigmentin hajaantuminen kaikkialle soluihin aiheuttaa tummentavan vaikutuksen. Pigmenttikäyttäytymistä ohjaavat hormonit, jotka tulevat silmänvarren neurosekretorisista soluista, samoin kuin verkkokalvon pigmentin siirtymistä silmien valo- ja pimeäadaptaatiota varten (kuva 19-8).
Neurosekretorisen aineen vapautuminen sydänpussin seinämässä olevista perikardiaalisista elimistä saa aikaan sydämen sykkeen nopeuden ja amplitudin lisääntymisen.
Androgeenisia rauhasia, jotka löydettiin ensimmäisenä amfipodista (Orchestia, yleinen rantahyppääjä), esiintyy urospuolisissa malakostraakissa. Toisin kuin useimmat muut äyriäisten endokriiniset elimet, ne eivät ole hermosekretorisia elimiä. Niiden eritys stimuloi uroksen sukupuoliominaisuuksien ilmentymistä. Nuorilla malacostracans-eläimillä on alkeelliset androgeeniset rauhaset, mutta naarailla nämä rauhaset eivät kehity. Jos ne istutetaan naaraaseen keinotekoisesti, sen munasarjat muuttuvat kiveksiksi ja alkavat tuottaa siittiöitä, ja sen lisäkkeet alkavat saada urospuolisia piirteitä seuraavassa moltissa. Isopodeilla androgeeniset rauhaset sijaitsevat kiveksissä; kaikilla muilla malacostracans-eläimillä ne ovat viimeisten rintakehän jalkojen coxopodien lihasten välissä ja osittain kiinnittyneinä vasa deferentian päiden läheisyyteen. Vaikka naarailla ei ole androgeenisten rauhasten kaltaisia elimiä, niiden munasarjat tuottavat yhtä tai kahta hormonia, jotka vaikuttavat toissijaisiin sukupuoliominaisuuksiin.
Hormoneja, jotka vaikuttavat muihin kehon prosesseihin, voi olla äyriäisissä, ja todisteet viittaavat siihen, että silmänvarressa tuotettu neurosekretorinen aine säätelee verensokerin tasoa.
Ruokailutottumukset
Ruokailutottumukset ja ruokailuun sopeutuminen vaihtelevat suuresti äyriäisten välillä. Monet muodot voivat siirtyä yhdestä ruokailutyypistä toiseen ympäristön ja ravinnon saatavuuden mukaan, mutta kaikki käyttävät samoja perustavanlaatuisia suupieliä. Alaleuat ja yläleuat toimivat ravinnon nielemiseen; yläleuat pitävät ja murskaavat ruokaa. Petoeläimillä kävelevät jalat, erityisesti sorkkajalat, palvelevat ravinnon pyydystämisessä.
Monet pienet ja suuret äyriäiset ovat petoeläimiä, ja joillakin niistä on mielenkiintoisia sopeutumia saaliin tappamiseen. Eräällä katkaravun kaltaisella muodolla, Lygiosquillalla, on yhdessä kävelevässä jalassaan erikoistunut sormi, joka voidaan vetää uraan ja vapauttaa äkillisesti lävistämään ohi kulkeva saalis. Pistoolikatkaravuilla (Alpheus spp.) on valtavan suuri kela, joka voidaan virittää kuin aseen vasara ja napsahtaa voimalla, joka tainnuttaa saaliin.
Suspensiosyöjien ravinto vaihtelee planktonista ja detrituksesta bakteereihin. Saalistajat syövät toukkia, matoja, äyriäisiä, etanoita ja kaloja. Aasinsyöjät syövät kuollutta eläin- ja kasviainesta. Riippusyöjät, kuten keijukatkaravut, vesikirput ja simpukat, käyttävät jalkojaan, joissa on paksut säikeet, luodakseen vesivirtauksia, jotka pyyhkivät ravintohiukkasia säikeiden läpi. Mutakatkaravut (Upogebia spp.) käyttävät kahden ensimmäisen parin rintakehän ulokkeidensa pitkiä ulokkeita siivilöidäkseen ruoka-ainesta vedestä, joka kiertää niiden kolon läpi niiden uikkareiden liikkeillä.
Ravuilla on kaksiosainen vatsa (kuva 19-13). Ensimmäisessä osassa on mahamylly, jossa alaleukojen jo repimä ruoka voidaan edelleen jauhaa kolmella kalkkihampaalla riittävän hienojakoisiksi hiukkasiksi, jotta ne voivat kulkea toisessa osassa olevan setoosisuodattimen läpi; ravintohiukkaset kulkeutuvat sitten suolistoon kemiallista ruoansulatusta varten.
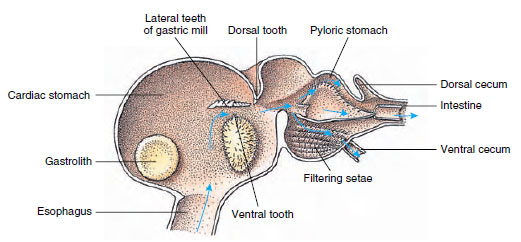
Kuva 19-13
Malacostracan mahalaukku, jossa näkyy mahalaukun ”mylly” ja ruoan liikesuunnat. Myllyssä on kitiiniharjanteita eli
hampaita pureskelua varten ja setaeja, jotka siivilöivät ruokaa ennen sen siirtymistä pyloriseen mahalaukkuun.
Vastaa