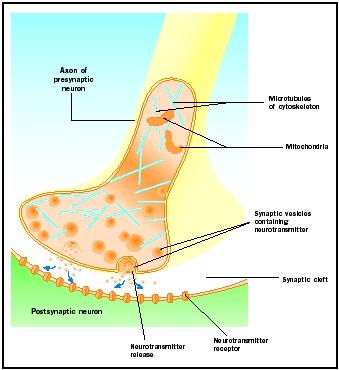
Liberación de neurotransmisores
Los potenciales de acción que llegan a los pomos sinápticos desencadenan la liberación de neurotransmisores en la hendidura sináptica. El mecanismo molecular no se conoce por completo. Entre la llegada del potencial de acción y la liberación del neurotransmisor se produce un «retraso sináptico» de uno a dos milisegundos. Los potenciales de acción abren canales de calcio en la membrana del pomo sináptico, lo que provoca un movimiento de entrada de iones de calcio . Los iones de calcio desencadenan la liberación del neurotransmisor desde las vesículas sinápticas hacia la hendidura sináptica. Las vesículas sinápticas se fusionan con la membrana presináptica durante este proceso de exocitosis. Las membranas de las vesículas antiguas pasan a formar parte de la membrana presináptica y las nuevas vesículas se desprenden de una zona adyacente de la membrana. Estas nuevas vesículas se rellenan posteriormente con neurotransmisores recién sintetizados o «reciclados» .
Los neurotransmisores liberados se difunden a través de la estrecha hendidura sináptica. En la membrana postsináptica, las moléculas de neurotransmisor se unen a moléculas receptoras unidas a la membrana con sitios de reconocimiento específicos para ese neurotransmisor. La unión del neurotransmisor al receptor desencadena una respuesta postsináptica específica para ese receptor. Estas respuestas pueden ser excitatorias o inhibitorias, dependiendo de las propiedades del receptor. Si la estimulación del receptor hace que la membrana postsináptica se vuelva eléctricamente más positiva (despolarizada), se trata de un potencial postsináptico excitador (EPSP). Si es más negativo (hiperpolarizado), se trata de un potencial postsináptico inhibitorio (IPSP). La excitación y la inhibición dependen de las propiedades del receptor y no del neurotransmisor. Los receptores acoplados a canales de sodio o de calcio son excitatorios y producen una despolarización de la membrana postsináptica, mientras que los receptores acoplados a canales de cloruro o de potasio son inhibitorios y producen una hiperpolarización de la membrana postsináptica. Estos receptores acoplados a canales iónicos se denominan receptores ionotrópicos.
Otros receptores están acoplados a sistemas de «segundo mensajero» que inician una serie de reacciones bioquímicas en la célula postsináptica. Estos son los receptores metabotrópicos . Los receptores metabotrópicos pueden producir muchas
eventos postsinápticos. Estos van desde la activación directa de los canales iónicos adyacentes, a la alteración de la sensibilidad del receptor, a la transcripción de ácidos ribonucleicos mensajeros (ARN) específicos, o incluso la activación de genes específicos . Las sinapsis químicas forman parte de un sistema de comunicación muy adaptable y flexible. No son estructuras anatómicas estáticas con propiedades fijas, sino que son estructuras dinámicas, capaces de cambiar sus propiedades moleculares con circunstancias cambiantes.
Hay literalmente cientos de neurotransmisores. Algunos son compuestos bastante simples como la acetilcolina, la serotonina, las catecolaminas (dopamina, norepinefrina y epinefrina) y varios de los aminoácidos . Muchos son más complejos y pertenecen a la amplia gama de neuropéptidos transmisores. Una vez liberados en la hendidura sináptica, los neurotransmisores permanecen activos hasta que son alterados químicamente o llevados de vuelta a la perilla sináptica por sistemas portadores especiales y reciclados. En las sinapsis colinérgicas, la acetilcolinesterasa está presente en la hendidura sináptica. Esta enzima escinde el neurotransmisor en acetato y colina, ninguno de los cuales es activo. La serotonina y la epinefrina, en cambio, son absorbidas por la terminal presináptica y recicladas.
Deja una respuesta