Subphylum Crustacea
Naturaleza general de un crustáceo
Los crustáceos se diferencian de otros artrópodos de diversas maneras, pero la única característica realmente distintiva es que los crustáceos son los únicos artrópodos con dos pares de antenas. Además de dos pares de antenas y un par de mandíbulas, los crustáceos tienen dos pares de maxilares en la cabeza, seguidos de un par de apéndices en cada segmento corporal o somito. En algunos crustáceos no todos los somitos llevan apéndices. Todos los apéndices, excepto quizás las primeras antenas, son primitivamente bífidos (dos ramas principales), y al menos algunos apéndices de los adultos actuales muestran esa condición. Los órganos especializados en la respiración, si están presentes, funcionan como branquias.
La mayoría de los crustáceos tienen entre 16 y 20 somitas, pero algunas formas tienen 60 somitas o más. Un mayor número de somitas es una característica primitiva. La condición más derivada es tener menos segmentos y una mayor tagmatización. Los principales tagmas son la cabeza, el tórax y el abdomen, pero éstos no son homólogos en toda la clase (o incluso dentro de algunas subclases) debido a los distintos grados de fusión de los somitas, por ejemplo, como en el cefalotórax.
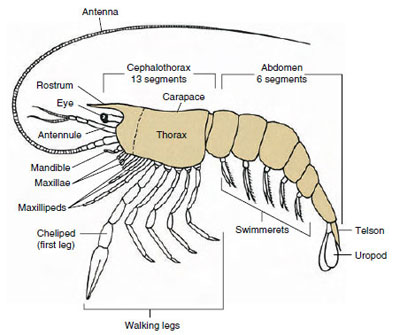
Figura 19-1
Plano arquetípico de Malacostraca. Los dos maxilares y los tres
maxilípedos se han separado en forma de diagrama para ilustrar el
plano general.
El grupo más numeroso de crustáceos es, con mucho, la clase Malacostraca, que incluye langostas, cangrejos, camarones, saltadores de playa, chinches y muchos otros. Estos muestran una disposición sorprendentemente constante de los segmentos corporales y los tagmata, que se considera el plan ancestral de la clase (Figura 19-1). Este plan corporal típico tiene una cabeza de cinco (seis embrionariamente) somitas fusionadas, un tórax de ocho somitas y un abdomen de seis somitas (siete en algunas especies). En el extremo anterior hay un rostrum no segmentado y en el extremo posterior hay un telson no segmentado, que con el último somito abdominal y sus urópodos forma un abanico de cola en muchas formas.
En muchos crustáceos la cutícula dorsal de la cabeza puede extenderse posteriormente y alrededor de los lados del animal para cubrir o estar fusionada con algunos o todos los somitos torácicos y abdominales. Esta cubierta se denomina caparazón. En algunos grupos, el caparazón forma válvulas en forma de concha que cubren la mayor parte o la totalidad del cuerpo. En los decápodos (incluyendo langostas, camarones, cangrejos y otros), el caparazón cubre todo el cefalotórax pero no el abdomen.
Forma y función
Debido a su tamaño y fácil disponibilidad, los crustáceos grandes como los cangrejos de río se han estudiado más que otros grupos. También son comúnmente estudiados en los cursos introductorios de laboratorio. Por lo tanto, muchos de los comentarios que siguen se aplican específicamente a los cangrejos de río y sus parientes.
Características externas
Los cuerpos de los crustáceos están cubiertos por una cutícula secretada compuesta de quitina, proteínas y material calcáreo. Las placas más duras y pesadas de los crustáceos de mayor tamaño están especialmente llenas de depósitos calcáreos. La dura cubierta protectora es blanda y fina en las uniones entre somitas, lo que permite la flexibilidad de movimiento. El caparazón, si está presente, cubre gran parte o la totalidad del cefalotórax; en los decápodos, como los cangrejos de río, todos los segmentos de la cabeza y el tórax están rodeados dorsalmente por el caparazón. Cada somito que no está encerrado en el caparazón está cubierto por una placa cuticular dorsal, o tergum (Figura 19-2A), y una barra transversal ventral, el esternón, se encuentra entre los apéndices segmentarios (Figura 19-2B). El abdomen termina en un telson, que no se considera un somito y lleva el ano. (El telson puede ser homólogo al pigidio de los anélidos).
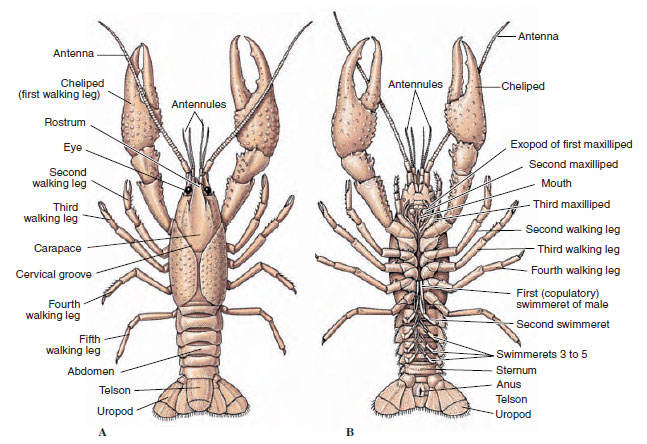
Figura 19-2
Estructura externa de los cangrejos de río. A, Vista dorsal. B, Vista ventral.
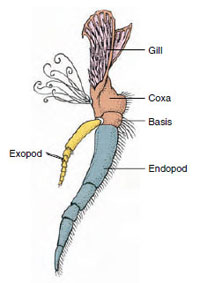
Figura 19-3
Partes de un apéndice de crustáceo bífido (tercer maxilipedo de un
cangrejo de río).
La posición de los gonoporos varía según el sexo y el grupo de crustáceos. Pueden estar sobre o en la base de un par de apéndices, en el extremo terminal del cuerpo o en somitas sin patas. En los cangrejos de río, las aberturas de los vasa deferentia se encuentran en el lado medio, en la base del quinto par de patas, y las de los oviductos en la base del tercer par. En las hembras, el orificio del receptáculo seminal suele estar situado en la línea media ventral entre el cuarto y el quinto par de patas caminadoras.
Apéndices: Los miembros de las clases Malacostraca (incluidos los cangrejos de río) y Remipedia suelen tener un par de apéndices articulados en cada somito (Figura 19-3), aunque los somitos abdominales de las demás clases no llevan apéndices. Los apéndices de los crustáceos derivados, como los cangrejos de río, presentan una considerable especialización. Sin embargo, todos son variaciones del plan básico bífido, ilustrado por un apéndice del cangrejo de río como el maxilipedo (una extremidad torácica modificada para convertirse en un apéndice de la cabeza) (Figuras 19-3 y 19-4). La porción basal, o protópodo, lleva un exópodo lateral y un endópodo medial. El protópodo está formado por una o dos articulaciones (base y coxa), mientras que el exópodo y el endópodo tienen de una a varias articulaciones cada uno. Algunos apéndices, como las patas para caminar de los cangrejos de río, son secundariamente unirrámeos. Las extremidades de los crustáceos presentan a veces procesos mediales o laterales, denominados endites y exites, respectivamente, y un exite en el protópodo se denomina epípodo. Los epípodos se modifican a menudo como branquias. La tabla 19-1 muestra cómo los distintos apéndices se han modificado a partir del plan birrámeo para adaptarse a funciones específicas.
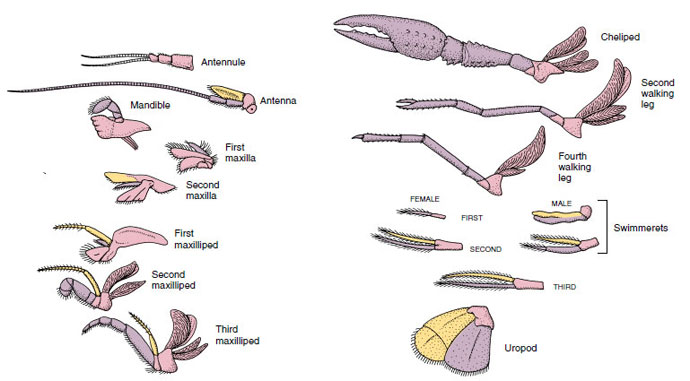
Figura 19-4
Apéndices de un cangrejo de río mostrando cómo se han modificado a partir del plan birrámeo básico, como se encuentra en un
swimmeret. Própodo, rosa; endópodo, púrpura; exópodo, amarillo.
Se dice que las estructuras que tienen un plan básico similar y que han descendido de una forma común son homólogas, tengan o no la misma función. Dado que las patas especializadas para caminar, las piezas bucales, los quelípedos y los nadadores se han desarrollado a partir de un tipo común de bípedo, pero se han modificado para realizar diferentes funciones, todos son homólogos entre sí, una condición conocida como homología en serie. Primitivamente las extremidades eran todas muy similares, pero durante la evolución de las modificaciones estructurales, algunas ramas se han reducido, otras se han perdido, otras se han alterado en gran medida y se han añadido algunas partes nuevas. Los cangrejos de río y sus aliados poseen la homología en serie más elaborada del reino animal, con 17 tipos de apéndices distintos pero homólogos en serie (Tabla 19-1).
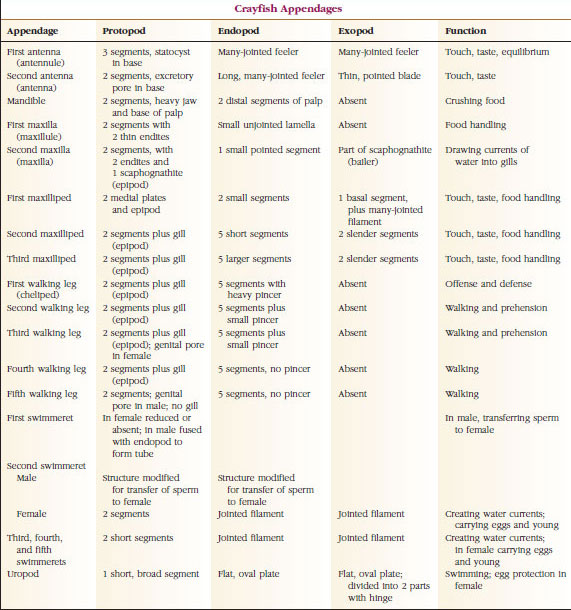
Tabla 19.1
Características internas
Los sistemas muscular y nervioso y la segmentación en el tórax y el abdomen muestran claramente el metamerismo heredado de los ancestros anélidos, pero hay marcadas modificaciones en otros sistemas. La mayoría de los cambios implican la concentración de partes en una región particular o bien la reducción o pérdida completa de partes, como el intersepta.
Hemocoel: El principal espacio corporal de los artrópodos no es el celoma sino un hemocoel lleno de sangre. Durante el desarrollo embrionario de la mayoría de los artrópodos, se abren cavidades celómicas vestigiales dentro del mesodermo de al menos algunos somitas. Pronto se obliteran o se vuelven continuas con el espacio entre las estructuras mesodérmicas y ectodérmicas en desarrollo y el vitelo. Este espacio se convierte en el hemocoel y, por tanto, no está revestido por un peritoneo mesodérmico. En los crustáceos los únicos compartimentos celómicos que quedan son los sacos terminales de los órganos excretores y el espacio alrededor de las gónadas.
Sistema muscular: Los músculos estriados constituyen una parte considerable del cuerpo de la mayoría de los crustáceos. Los músculos suelen estar dispuestos en grupos antagónicos: flexores, que atraen una parte hacia el cuerpo, y extensores, que la extienden hacia fuera. El abdomen de un cangrejo de río tiene potentes flexores (Figura 19-5), que se utilizan cuando el animal nada hacia atrás, su mejor medio de escape. Unos fuertes músculos situados a ambos lados del estómago controlan las mandíbulas.
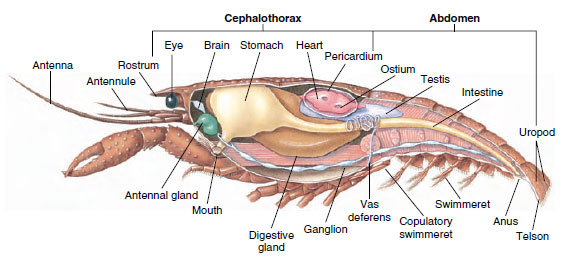
Figura 19-5
Estructura interna de un cangrejo de río macho.
Sistema respiratorio: El intercambio de gases respiratorios en los crustáceos más pequeños se produce en zonas más delgadas de la cutícula (por ejemplo, en las patas) o en todo el cuerpo, y las estructuras especializadas pueden estar ausentes. Los crustáceos más grandes tienen branquias, que son delicadas proyecciones en forma de pluma con una cutícula muy fina. En los decápodos, los lados del caparazón encierran la cavidad branquial, que está abierta anterior y ventralmente (Figura 19-6). Las branquias pueden proyectarse desde la pared pleural hacia la cavidad branquial, desde la articulación de las patas torácicas con el cuerpo o desde las coxas torácicas. Estos dos últimos tipos son típicos de los cangrejos de río. El «achicador», una parte del segundo maxilar, arrastra el agua por encima de los filamentos branquiales, hacia la cavidad branquial en las bases de las patas, y hacia fuera de la cavidad branquial en la parte anterior.
Sistema circulatorio: Los crustáceos y otros artrópodos tienen un sistema circulatorio de tipo «abierto» o lacunar. Esto significa que no hay venas ni separación de la sangre del líquido intersticial, como ocurre en los animales con sistemas cerrados. La hemolinfa (sangre) sale del corazón a través de las arterias, circula por el hemocele y vuelve a los senos venosos, o espacios, en lugar de las venas, antes de volver a entrar en el corazón. Los anélidos tienen un sistema cerrado, al igual que los vertebrados.
El corazón dorsal es el principal órgano propulsor. Es un saco unicameral de músculo estriado. La hemolinfa entra en el corazón desde el seno pericárdico que lo rodea a través de ostias pareadas, con válvulas que impiden el reflujo hacia el seno (Figura 19-6). Desde el corazón, la hemolinfa entra en una o más arterias. Las válvulas de las arterias impiden el reflujo de la hemolinfa. Las arterias pequeñas desembocan en los senos tisulares, que a su vez suelen descargar en un gran seno esternal (Figura 19-6).
Desde allí, los canales aferentes de los senos llevan la hemolinfa a las branquias, si están presentes, para el intercambio de oxígeno y dióxido de carbono. La hemolinfa vuelve entonces al seno pericárdico por los canales eferentes (Figura 19-6).
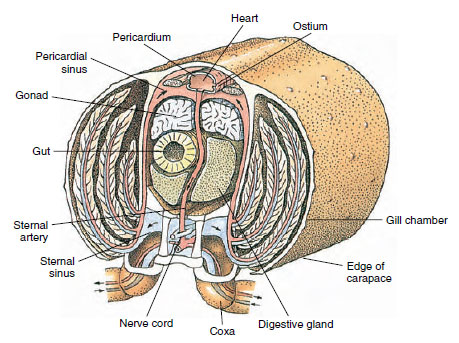
Figura 19-6
Sección transversal diagramática a través de la región del corazón de un cangrejo de río que muestra la dirección del flujo sanguíneo en este sistema sanguíneo «abierto»
. El corazón bombea sangre a los tejidos del cuerpo a través de las arterias, que desembocan en los senos tisulares. La sangre que regresa entra en el seno esternal, luego pasa por las branquias para el intercambio de gases y finalmente vuelve al seno pericárdico por los canales eferentes. Obsérvese la ausencia de venas.
La hemolinfa en los artrópodos es en gran parte incolora. Incluye células ameboides de al menos dos tipos. La hemocianina, un pigmento respiratorio que contiene cobre, o la hemoglobina, un pigmento que contiene hierro, pueden ser transportados en solución. La hemolinfa tiene la propiedad de coagularse, lo que evita su pérdida en lesiones menores. Algunas células ameboides liberan un coagulante similar a la trombina que precipita la coagulación.
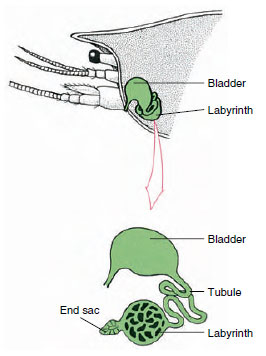
Figura 19-7
Esquema de la glándula antenal (glándula verde) de
los cangrejos de río. (En posición natural el órgano está muy
plegado.) Algunos crustáceos carecen de laberinto,
y el túbulo excretor (canal nefridial) es
un tubo muy enrollado.
Sistema excretor: Los órganos excretores de los crustáceos adultos son un par de estructuras tubulares situadas en la parte ventral de la cabeza por delante del esófago (Figura 19-5). Se denominan glándulas antenales o glándulas maxilares, según se abran en la base de las antenas o del segundo maxilar. Algunos crustáceos adultos tienen ambas. Los órganos excretores de los decápodos son las glándulas antenales, también llamadas glándulas verdes en este grupo. Los crustáceos no tienen túbulos malpighianos, los órganos excretores de las arañas y los insectos.
El saco final de la glándula antenal, que deriva de un compartimento coelómico embrionario, consta de una pequeña vesícula (sáculo) y una masa esponjosa llamada laberinto. El laberinto se conecta mediante un túbulo excretor a una vejiga dorsal, que se abre al exterior por un poro en la superficie ventral del segmento antenal basal (Figura 19-7). La presión hidrostática dentro de la hemocoela proporciona la fuerza para la filtración del líquido en el saco terminal. A medida que el filtrado pasa por el túbulo excretor y la vejiga, se modifica mediante la reabsorción de sales, aminoácidos, glucosa y algo de agua, y finalmente se excreta en forma de orina.
La excreción de desechos nitrogenados (sobre todo amoníaco) tiene lugar por difusión a través de zonas finas de la cutícula, especialmente las branquias, y los llamados órganos excretores funcionan principalmente para regular la composición iónica y osmótica de los fluidos corporales. Los crustáceos de agua dulce, como los cangrejos de río, se ven constantemente amenazados por la sobredilución de su sangre por el agua, que se difunde a través de las branquias y otras superficies permeables al agua. Las glándulas verdes, al formar una orina diluida y poco salada, actúan como un eficaz dispositivo de «control de inundaciones». En la orina se pierde algo de Na+ y Cl-, pero esta pérdida se compensa con la absorción activa de sal disuelta por las branquias. En los crustáceos marinos, como las langostas y los cangrejos, el riñón funciona para ajustar la composición de sal de la hemolinfa mediante la modificación selectiva del contenido de sal de la orina tubular. En estas formas, la orina permanece isosmótica con respecto a la sangre.
Sistemas nervioso y sensorial: Los sistemas nerviosos de los crustáceos y los anélidos tienen mucho en común, aunque los de los crustáceos tienen más fusión de ganglios (Figura 19-5). El cerebro es un par de ganglios supraesofágicos que suministran nervios a los ojos y a dos pares de antenas. Está unido por conectivos al ganglio subesofágico, una fusión de al menos cinco pares de ganglios que suministran nervios a la boca, los apéndices, el esófago y las glándulas antenales. El doble cordón nervioso ventral tiene un par de ganglios para cada somita y nervios que sirven a los apéndices, los músculos y otras partes.
Además de este sistema central, puede haber un sistema nervioso simpático asociado al tracto digestivo.
Los crustáceos tienen órganos sensoriales mejor desarrollados que los anélidos. Los mayores órganos sensoriales de los cangrejos de río son los ojos y los estatocistos. Los pelos táctiles, delicadas proyecciones de la cutícula, están ampliamente distribuidos por el cuerpo y son especialmente abundantes en los quelos, las piezas bucales y el telson. Los sentidos químicos del gusto y el olfato se encuentran en los pelos de las antenas, las piezas bucales y otros lugares.
En el segmento basal de cada primera antena de los cangrejos de río se encuentra un estatocisto en forma de saco que se abre a la superficie por un poro dorsal. El estatocisto contiene una cresta que lleva pelos sensoriales formados por el revestimiento quitinoso y granos de arena que sirven de estatolitos. Cada vez que el animal cambia de posición, los cambios correspondientes en la posición de los granos de los pelos sensoriales se transmiten como estímulos al cerebro, y el animal puede ajustarse en consecuencia. Con cada muda (ecdisis) de la cutícula se pierde el revestimiento cuticular del estatocisto y con él los granos de arena. Después de la ecdisis se recogen nuevos granos a través del poro dorsal.
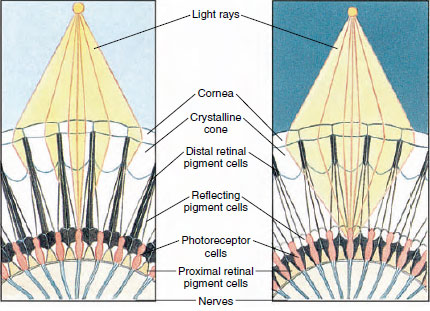
Figura 19-8
Porción del ojo compuesto de un artrópodo que muestra la migración del pigmento
en los ommatidios para la visión diurna y nocturna. Se representan cinco ommatidios en
cada diagrama. Durante el día, cada ommatidio está rodeado por un collar de
pigmento oscuro, de modo que cada ommatidio es estimulado sólo por los rayos de luz
que entran en su propia córnea (visión en mosaico); durante la noche, el pigmento forma collares
incompletos y los rayos de luz pueden extenderse a los ommatidios adyacentes
(imagen continua, o de superposición).
Los ojos de muchos crustáceos son compuestos, formados por muchas unidades fotorreceptoras llamadas ommatidios (Figura 19-8). Cubriendo la superficie redondeada de cada ojo hay una zona transparente de cutícula, la córnea, que está dividida en muchos pequeños cuadrados o hexágonos conocidos como facetas. Estas facetas son los extremos exteriores de los ommatidios. Cada ommatidio se comporta como un ojo diminuto y contiene varios tipos de células dispuestas en forma de columna (Figura 19-8). Las células pigmentarias negras se encuentran entre los ommatidios adyacentes.
El movimiento del pigmento en un ojo compuesto de artrópodo le permite ajustarse a diferentes cantidades de luz. Hay tres conjuntos de células pigmentarias en cada ommatidio: retina distal, retina proximal y reflectante; éstas están dispuestas de tal manera que pueden formar un collar o manguito más o menos completo alrededor de cada ommatidio. En caso de luz intensa o de adaptación diurna, el pigmento retiniano distal se desplaza hacia dentro y se encuentra con el pigmento retiniano proximal, que se desplaza hacia fuera, de modo que se forma un manguito pigmentario completo alrededor del ommatidio (figura 19-8). En este estado, sólo los rayos que inciden directamente en la córnea alcanzan las células fotorreceptoras (retinulares), ya que cada omatidios está protegido de los demás. Por lo tanto, cada ommatidium verá sólo una zona limitada del campo de visión (una imagen en mosaico, o de aposición). Con luz tenue, los pigmentos distales y proximales se separan para que los rayos de luz, con la ayuda de las células pigmentarias reflectantes, tengan la oportunidad de propagarse a los omatidios adyacentes y formar una imagen continua o de superposición. Este segundo tipo de visión es menos preciso, pero aprovecha al máximo la cantidad limitada de luz recibida.
Reproducción, ciclos vitales y función endocrina
La mayoría de los crustáceos tienen sexos separados, y existen varias especializaciones para la cópula entre los distintos grupos. Los percebes son monoicos pero generalmente practican la fecundación cruzada. En algunos ostrácodos los machos son escasos y la reproducción suele ser partenogenética. La mayoría de los crustáceos empollan sus huevos de alguna manera: los branquiópodos y los percebes tienen cámaras de cría especiales, los copépodos tienen sacos de cría adheridos a los lados del abdomen (véase la figura 19-19), y muchos malacostracos llevan huevos y crías adheridos a sus apéndices abdominales.
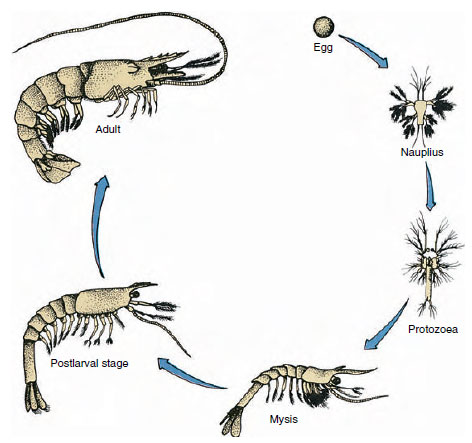
Figura 19-9
Ciclo vital de un camarón del Golfo Penaeus. Los peneidos desovan a profundidades de 40 a 90 m. Las formas larvarias jóvenes son
planctónicas y se desplazan hacia la costa a aguas de menor salinidad para desarrollarse como juveniles. Los camarones más viejos regresan a aguas más profundas
en alta mar.
Los cangrejos de río tienen un desarrollo directo: no hay forma larvaria. Del huevo nace un diminuto juvenil con la misma forma que el adulto y un conjunto completo de apéndices y somitas. Sin embargo, el desarrollo es indirecto en la mayoría de los crustáceos, y del huevo nace una larva muy diferente al adulto en cuanto a estructura y aspecto. El cambio de larva a adulto es la metamorfosis. La larva primitiva y más extendida en los crustáceos es el nauplio (figuras 19-9 y 19-23). Los nauplios sólo tienen tres pares de apéndices: las primeras anténulas parciales, las antenas parciales y las mandíbulas parciales. Todos funcionan como apéndices natatorios en esta fase.
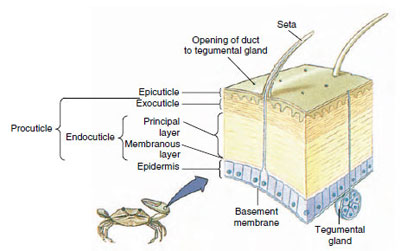
Figura 19-10
Estructura de la cutícula de los crustáceos.
El desarrollo posterior puede implicar un cambio gradual hacia la forma corporal adulta, y los apéndices y somitas se añaden a través de una serie de mudas, o la asunción de la forma adulta puede implicar cambios más bruscos. Por ejemplo, la metamorfosis de un percebe pasa de ser un nauplio que nada libremente a una larva con un caparazón bivalvo llamado ciprés y finalmente a un adulto sésil con placas calcáreas.
Ecdisis: La ecdisis (ek’duh-sis) (Gr. ekdyein, despojarse), o muda, es necesaria para que el cuerpo aumente de tamaño porque el exoesqueleto no es vivo y no crece a medida que el animal crece. Gran parte del funcionamiento de un crustáceo, incluyendo su reproducción, comportamiento y muchos procesos metabólicos, se ve directamente afectado por la fisiología del ciclo de muda.
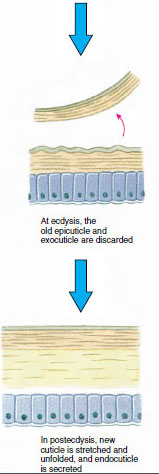
La cutícula, que es secretada por la epidermis subyacente, tiene varias capas (Figura 19-10). La más externa es la epicutícula, una capa muy fina de proteína impregnada de lípidos. El grueso de la cutícula son las diversas capas de la procuticula: (1) la exocutícula, que se encuentra justo debajo de la epicutícula y contiene proteínas, sales de calcio y quitina; (2) la endocutícula, que a su vez se compone de (3) una capa principal, que contiene más quitina y menos proteínas y está fuertemente calcificada, y (4) una capa membranosa no calcificada, una capa relativamente fina de quitina y proteínas.
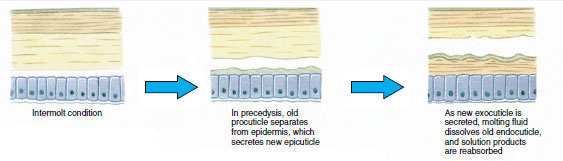
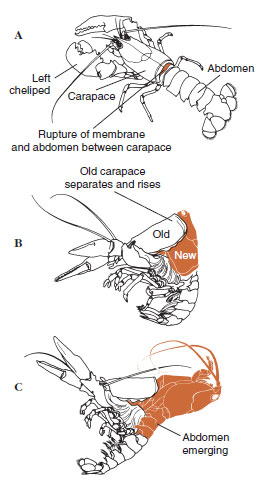
Un tiempo antes de la ecdisis real, las células epidérmicas se agrandan considerablemente. Se separan de la capa membranosa, segregan una nueva epicutícula y comienzan a segregar una nueva exocutícula (Figura 19-11). Se liberan enzimas en la zona por encima de la nueva epicutícula. Estas enzimas comienzan a disolver la vieja endocutícula, y los productos solubles son reabsorbidos y almacenados dentro del cuerpo del crustáceo. Algunas sales de calcio se almacenan como gastrolitos (acumulaciones minerales) en las paredes del estómago. Finalmente, sólo quedan la exocutícula y la epicutícula de la antigua cutícula, recubiertas por una nueva epicutícula y una nueva exocutícula. El animal traga agua, que absorbe a través del intestino, y su volumen de sangre aumenta considerablemente. La presión interna hace que la cutícula se parta y el animal se desprende de su antiguo exoesqueleto (Figura 19-12). A continuación se produce un estiramiento de la nueva cutícula, aún blanda, la deposición de la nueva endocuticula, la redeposición de las sales inorgánicas recuperadas y otros componentes, y el endurecimiento de la nueva cutícula. Durante el periodo de muda, el animal está indefenso y permanece oculto.
 |
|
 |
 |
| Figura 19-12 Secuencia de muda en una langosta, Homarus americanus. A, La membrana entre el caparazón y el abdomen se rompe, y el caparazón comienza a elevarse lentamente. Este paso puede durar hasta 2 horas. B y C, La cabeza, el tórax y finalmente el abdomen se retiran. Este proceso no suele durar más de 15 minutos. Inmediatamente después de la ecdisis, los quelípedos están desecados y el cuerpo es muy blando. La langosta continúa con la rápida absorción de agua, de modo que en 12 horas el cuerpo aumenta aproximadamente un 20% en longitud y un 50% en peso. El agua de los tejidos será reemplazada por proteínas en las semanas siguientes. |
Figura 19-11 Secreción de cutículas y resorción en la ecdisis. |
Cuando un crustáceo es joven, la ecdisis debe producirse con frecuencia para permitir el crecimiento, y el ciclo de muda es relativamente corto. A medida que el animal se acerca a la madurez, los periodos de muda se alargan progresivamente y, en algunas especies, la muda cesa por completo. Durante los periodos de intermutación, se produce un aumento de la masa tisular a medida que el tejido vivo sustituye al agua.
Control hormonal del ciclo de ecdisis
Aunque la ecdisis está controlada hormonalmente, el ciclo suele iniciarse por un estímulo ambiental percibido por el sistema nervioso central. Estos estímulos pueden incluir la temperatura, la duración del día y la humedad (en el caso de los cangrejos terrestres). La señal del sistema nervioso central disminuye la producción de una hormona inhibidora de la muda por parte del órgano X. El órgano X es un grupo de células neurosecretoras en la médula terminal del cerebro. En los cangrejos de río y otros decápodos, la médula terminal se encuentra en el pedúnculo ocular. La hormona es transportada por los axones del órgano X hasta la glándula sinusal (que probablemente no tenga una función glandular), también en el pedúnculo ocular, donde se libera en la hemolinfa.
Un descenso en el nivel de la hormona inhibidora de la muda promueve la liberación de una hormona de la muda desde los órganos Y. Los órganos Y se encuentran bajo la epidermis, cerca de los músculos aductores de las mandíbulas, y son homólogos a las glándulas protorácicas de los insectos, que producen la hormona ecdisona. La acción de la hormona de la muda es iniciar los procesos que conducen a la ecdisis (proecdisis). Una vez iniciado, el ciclo procede automáticamente sin más acción de las hormonas de los órganos X o Y.
Otras funciones endocrinas: No sólo la eliminación de los pedúnculos oculares acelera la muda, sino que también se descubrió hace más de 100 años que los crustáceos a los que se les han eliminado los pedúnculos oculares ya no pueden ajustar la coloración del cuerpo a las condiciones de fondo. Hace más de 50 años se descubrió que el defecto no estaba causado por la pérdida de la visión, sino por la pérdida de hormonas en los oculares. El color del cuerpo de los crustáceos es, en gran medida, el resultado de los pigmentos de las células ramificadas especiales (cromatóforos) de la epidermis.
La concentración de gránulos de pigmento en el centro de las células provoca un efecto de aclaración, y la dispersión del pigmento por todas las células provoca un efecto de oscurecimiento. El comportamiento del pigmento está controlado por las hormonas de las células neurosecretoras del pedúnculo ocular, al igual que la migración del pigmento de la retina para la adaptación a la luz y a la oscuridad en los ojos (Figura 19-8).
La liberación de material neurosecretor de los órganos pericárdicos en la pared del pericardio provoca un aumento de la frecuencia y la amplitud de los latidos del corazón.
Las glándulas andrógenas, que se encontraron por primera vez en un anfípodo (Orchestia, un saltamontes de playa común), se dan en los malacostracos macho. A diferencia de la mayoría de los órganos endocrinos de los crustáceos, no son órganos neurosecretores. Su secreción estimula la expresión de los caracteres sexuales masculinos. Los malacostracos jóvenes tienen glándulas andrógenas rudimentarias, pero en las hembras estas glándulas no se desarrollan. Si se implantan artificialmente en una hembra, sus ovarios se transforman en testículos y comienzan a producir esperma, y sus apéndices comienzan a adoptar características masculinas en la siguiente muda. En los isópodos las glándulas andrógenas se encuentran en los testículos; en todos los demás malacostracos están entre los músculos de los coxópodos de las últimas patas torácicas y parcialmente unidas cerca de los extremos de los vasa deferentia. Aunque las hembras no poseen órganos similares a las glándulas andrógenas, sus ovarios producen una o dos hormonas que influyen en los caracteres sexuales secundarios.
Puede haber hormonas que influyan en otros procesos corporales de los crustáceos, y las pruebas sugieren que una sustancia neurosecretoria producida en el pedúnculo ocular regula el nivel de azúcar en la sangre.
Hábitos alimenticios
Los hábitos alimenticios y las adaptaciones para la alimentación varían mucho entre los crustáceos. Muchas formas pueden cambiar de un tipo de alimentación a otro dependiendo del entorno y de la disponibilidad de alimentos, pero todos utilizan el mismo conjunto fundamental de piezas bucales. Las mandíbulas y los maxilares sirven para ingerir el alimento; los maxilípedos sujetan y trituran el alimento. En los depredadores, las patas que caminan, especialmente los quelípedos, sirven para capturar el alimento.
Muchos crustáceos, tanto grandes como pequeños, son depredadores, y algunos tienen interesantes adaptaciones para matar a sus presas. Una forma parecida a la gamba, la Lygiosquilla, tiene en una de sus patas un dedo especializado que puede introducirse en una ranura y soltarse repentinamente para perforar a la presa que pasa. Los camarones pistola (Alpheus spp.) tienen una quela enormemente agrandada que puede amartillarse como el martillo de una pistola y romperse con una fuerza que aturde a su presa.
El alimento de los alimentadores en suspensión va desde el plancton y los detritus hasta las bacterias. Los depredadores consumen larvas, gusanos, crustáceos, caracoles y peces. Los carroñeros se alimentan de materia animal y vegetal muerta. Los que se alimentan en suspensión, como los camarones hada, las pulgas de agua y los percebes, utilizan sus patas, que llevan una gruesa franja de setas, para crear corrientes de agua que arrastran las partículas de alimento a través de las setas. Los camarones del fango (Upogebia spp.) utilizan largas setas en sus dos primeros pares de apéndices torácicos para colar el material alimenticio del agua que circula por su madriguera mediante los movimientos de sus nadadores.
Los cangrejos de río tienen un estómago dividido en dos partes (Figura 19-13). La primera parte contiene un molino gástrico en el que el alimento, ya desgarrado por las mandíbulas, puede ser triturado aún más por tres dientes calcáreos en partículas lo suficientemente finas como para pasar a través de un filtro de setas en la segunda parte; las partículas de alimento pasan entonces al intestino para la digestión química.
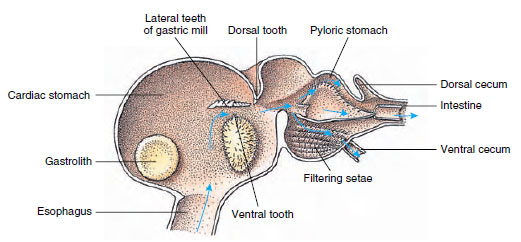
Figura 19-13
Estómago de Malacostra mostrando el «molino» gástrico y las direcciones de los movimientos del alimento. El molino tiene crestas quitinosas, o
dientes, para la masticación, y setas para colar el alimento antes de que pase al estómago pilórico.
Deja una respuesta