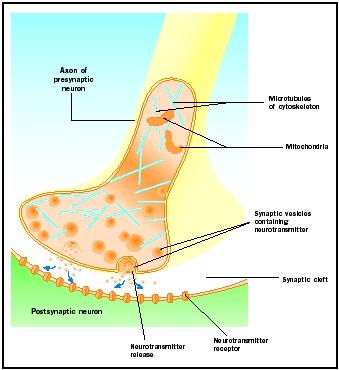
Frigivelse af neurotransmitter
Aktionspotentialer, der ankommer til synaptiske knopper, udløser frigivelse af neurotransmitter i den synaptiske kløft. Den molekylære mekanisme er ikke helt forstået. Der opstår en “synaptisk forsinkelse” på et til to millisekunder mellem ankomsten af aktionspotentialet og neurotransmitterfrigivelsen. Aktionspotentialer åbner calciumkanaler i den synaptiske knops membran, hvilket medfører en indadgående bevægelse af calciumioner . Calciumionerne udløser frigivelsen af neurotransmitter fra synaptiske vesikler i den synaptiske kløft. De synaptiske vesikler smelter sammen med den præsynaptiske membran under denne eksocytoseproces. Membranerne fra de gamle vesikler bliver en del af den præsynaptiske membran, og nye vesikler klemmes af fra et tilstødende område af membranen. Disse nye vesikler fyldes efterfølgende igen med nysyntetiserede eller “genbrugte” neurotransmittere .
De frigjorte neurotransmittere diffunderer gennem den smalle synaptiske kløft. Ved den postsynaptiske membran binder neurotransmittermolekyler sig til membranbundne receptormolekyler med genkendelsessteder, der er specifikke for den pågældende neurotransmitter. Neurotransmitterens binding til receptoren udløser et postsynaptisk respons, der er specifikt for den pågældende receptor. Disse reaktioner kan være enten excitatoriske eller hæmmende, afhængigt af receptorens egenskaber. Hvis stimulering af receptoren resulterer i, at den postsynaptiske membran bliver mere elektrisk positiv (depolariseret), er der tale om et excitatorisk postsynaptisk potentiale (EPSP). Hvis den bliver mere negativ (hyperpolariseret), er der tale om et hæmmende postsynaptisk potentiale (IPSP). Excitation og hæmning afhænger af receptorens egenskaber og ikke af neurotransmitteren. Receptorer, der er koblet til natrium- eller calciumkanaler, er excitatoriske og giver en depolarisering af den postsynaptiske membran, mens receptorer, der er koblet til klorid- eller kaliumkanaler, er hæmmende og giver en hyperpolarisering af den postsynaptiske membran. Sådanne receptorer, der er koblet til ionkanaler, kaldes ionotrope receptorer.
Andre receptorer er koblet til “second-messenger”-systemer, der igangsætter en række biokemiske reaktioner i den postsynaptiske celle. Disse er metabotropiske receptorer . Metabotrope receptorer kan producere mange forskellige
postsynaptiske hændelser. Disse spænder fra direkte aktivering af tilstødende ionkanaler til ændring af receptorfølsomhed, til transkription af specifikke budbringerribonukleinsyrer (RNA’er) eller endog aktivering af specifikke gener . Kemiske synapser er en del af et meget tilpasningsdygtigt og fleksibelt kommunikationssystem. Der er ikke tale om statiske anatomiske strukturer med faste egenskaber, men om dynamiske strukturer, der er i stand til at ændre deres molekylære egenskaber under skiftende omstændigheder.
Der findes bogstaveligt talt hundredvis af neurotransmittere. Nogle er ret enkle forbindelser som f.eks. acetylcholin, serotonin, katekolaminerne (dopamin, noradrenalin og adrenalin) og en række af aminosyrerne . Mange er mere komplekse og hører til det store udvalg af neuropeptidtransmittere. Når de først er frigivet i den synaptiske kløft, forbliver neurotransmittere aktive, indtil de enten ændres kemisk eller tages tilbage i den synaptiske knude af særlige transportsystemer og genbruges. Ved kolinerge synapser findes acetylcholinesterase i den synaptiske kløft. Dette enzym spalter neurotransmitteren til acetat og cholin, som begge ikke er aktive. Serotonin og adrenalin optages derimod i den præsynaptiske terminal og genanvendes.
Skriv et svar