Subphylum Crustacea
Generel karakter af et krebsdyr
Krebsdyr adskiller sig fra andre leddyr på mange forskellige måder, men det eneste kendetegn, der virkelig adskiller dem fra andre leddyr, er, at krebsdyr er de eneste leddyr, der har to par antenner. Ud over to par antenner og et par underkæber har krebsdyr to par overkæber på hovedet og derefter et par vedhæng på hvert kropssegment eller somit. Hos nogle krebsdyr er det ikke alle somitter, der er forsynet med vedhæng. Alle vedhæng, undtagen måske de første antenner, er oprindeligt biramøse (to hovedgrene), og i det mindste nogle af vedhængene hos nutidens voksne dyr viser denne tilstand. Organer, der er specialiseret til åndedræt, fungerer som gæller, hvis de er til stede.
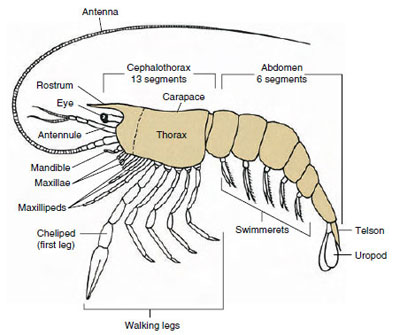
De fleste krebsdyr har mellem 16 og 20 somitter, men nogle former har 60 somitter eller mere. Et større antal somitter er et primitivt træk. Den mere afledte tilstand er at have færre segmenter og øget tagmatisering. De vigtigste tagmata er hoved, thorax og abdomen, men disse er ikke homologe i hele klassen (eller endog inden for nogle underklasser) på grund af forskellige grader af sammensmeltning af somitter, f.eks. som i cephalothorax.
Figur 19-1
Arkettypisk plan af Malacostraca. De to maxillae og tre
maxillipeds er skematisk adskilt for at illustrere den
generelle plan.
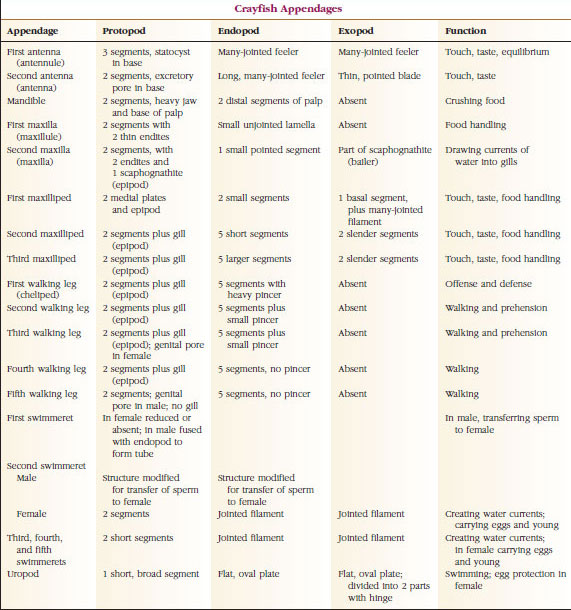
Langt den største gruppe af krebsdyr er klassen Malacostraca, som omfatter hummere, krabber, rejer, strandhopper, sokseorme og mange andre. Disse viser et overraskende konstant arrangement af kropssegmenter og tagmata, som anses for at være klassens forfødte plan (Figur 19-1). Denne typiske kropsplan har et hoved af fem (seks embryonalt) sammensmeltede somitter, et thorax af otte somitter og et abdomen af seks somitter (syv hos nogle få arter). I den forreste ende er der et ikke-segmenteret rostrum, og i den bageste ende er der et ikke-segmenteret telson, som sammen med den sidste abdominalsomit og dens uropoder danner en halefane hos mange former.
I mange krebsdyr kan hovedets dorsalcutikula strække sig bagud og rundt om dyrets sider for at dække eller være smeltet sammen med nogle af eller alle thorakal- og abdominalsomitterne. Denne beklædning kaldes en carapace. Hos nogle grupper danner carapacen muslingeskallignende ventiler, der dækker det meste af eller hele kroppen. Hos decapoder (bl.a. hummere, rejer og krabber) dækker panseret hele cephalothorax, men ikke maven.
Form og funktion
På grund af deres størrelse og lette tilgængelighed er store krebsdyr som f.eks. krebsdyr blevet undersøgt mere end andre grupper. De er også almindeligt studeret i indledende laboratoriekurser. Derfor gælder mange af de følgende kommentarer specifikt for krebs og deres slægtninge.
Udvendige kendetegn
Krebsdyrs kroppe er dækket af en udskilt kutikula bestående af chitin, protein og kalkholdigt materiale. De større krebsdyrs hårdere, tunge plader har et særligt højt indhold af kalkholdige aflejringer. Den hårde beskyttende belægning er blød og tynd ved leddene mellem somitterne, hvilket giver mulighed for fleksibilitet i bevægelserne. Carapax, hvis den er til stede, dækker en stor del af eller hele cephalothorax; hos decapoder som f.eks. krebsdyr er alle hoved- og brystsegmenter omsluttet dorsalt af carapax. Hver somit, der ikke er omsluttet af carapace, er dækket af en dorsal cuticularplade eller tergum (Figur 19-2A), og en ventral tværgående stang, sternum, ligger mellem segmenttilhængene (Figur 19-2B). Bagkroppen afsluttes med et telson, som ikke betragtes som en somit og bærer anus. (telsonet kan være homologt til annelidernes pygidium.)
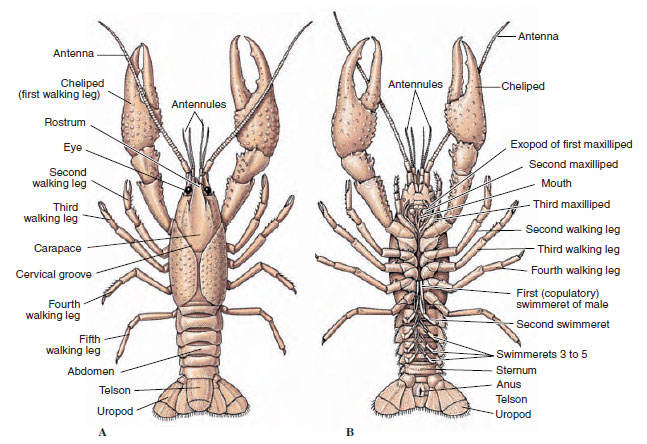
Figur 19-2
Krebsers ydre opbygning. A, Dorsal set. B, Ventral view.
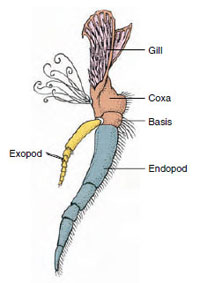
Figur 19-3
Dele af et biramøst krebsdyr
tilbehør (tredje maxilliped af en
krebs).
Gonoporernes placering varierer alt efter køn og gruppe af krebsdyr. De kan sidde på eller ved basis af et par vedhæng, i den afsluttende ende af kroppen eller på somitter uden ben. Hos krebsene er åbningerne til vasa deferentia på mediansiden ved basis af det femte par gående ben, og ovidukternes åbninger er ved basis af det tredje par. Hos hunnerne er åbningen til sædbeholderen normalt placeret i den midventrale linje mellem det fjerde og femte par af gangbenene.
Anlægsorganer: Medlemmer af klasserne Malacostraca (herunder krebs) og Remipedia har typisk et par leddelte vedhæng på hver somit (Figur 19-3), selv om de abdominale somitter i de andre klasser ikke bærer vedhæng. En betydelig specialisering er tydelig i vedhæng hos afledte krebsdyr som f.eks. krebsdyr. Alle er dog variationer af den grundlæggende, biramøse plan, som illustreres af et krebsevedhæng som f.eks. et maxilliped (et brystben, der er modificeret til at blive et hovedvedhæng) (figur 19-3 og 19-4). Den basale del, eller protopoden, har en lateral exopode og en medial endopode. Protopoden består af et eller to led (basis og coxa), mens exopoden og endopoden hver har fra et til flere led. Nogle vedhæng, f.eks. vandreben hos krebs, er sekundært uniramøse. Der forekommer undertiden mediale eller laterale processer på krebsdyrs lemmer, som kaldes henholdsvis endites og exites, og en exite på protopoden kaldes en epipode. Epipoder er ofte modificeret som gæller. Tabel 19-1 viser, hvordan de forskellige vedhæng er blevet modificeret fra den biramøse plan for at passe til specifikke funktioner.
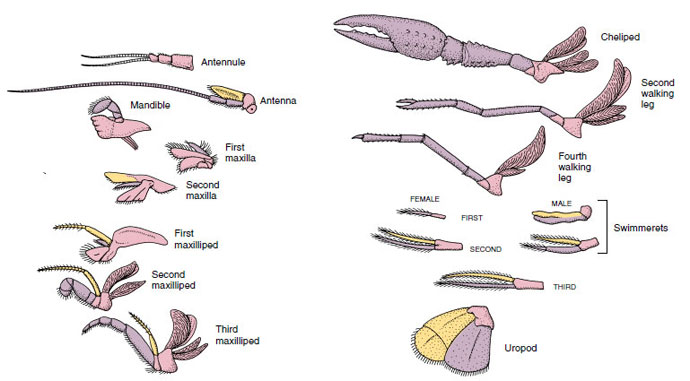
Figur 19-4
Anlægsorganer hos en flodkrebs, der viser, hvordan de er blevet modificeret fra den grundlæggende biramøse plan, som det findes hos en
svømmeret. Protopod, lyserød; endopod, lilla; exopod, gul.
Strukturer, der har en lignende grundplan og er nedstammet fra en fælles form, siges at være homologe, uanset om de har samme funktion eller ej. Da specialiserede gangben, munddele, chelipeder og svømmefødder alle har udviklet sig ud fra en fælles biramøs type, men er blevet modificeret til at udføre forskellige funktioner, er de alle homologe i forhold til hinanden, en tilstand, der kaldes seriel homologi. Primitivt var lemmerne alle meget ens, men i løbet af udviklingen af strukturelle modifikationer er nogle grene blevet reduceret, nogle gået tabt, nogle kraftigt ændret, og nogle nye dele er blevet tilføjet. Flodkrebs og deres allierede besidder den mest udførlige serielle homologi i dyreriget, idet de har 17 forskellige, men serielt homologe typer af vedhæng (tabel 19-1).
TABEL 19.1
Interne træk
Muskel- og nervesystemet og segmenteringen i thorax og abdomen viser tydeligt metamerisme, der er nedarvet fra annelide-lignende forfædre, men der er markante modifikationer i andre systemer. De fleste ændringer indebærer en koncentration af dele i et bestemt område eller en reduktion eller et fuldstændigt tab af dele, som f.eks. intersepta.
Hemocoel: Det største kropsrum hos leddyrene er ikke coelom, men et blodfyldt hemocoel. Under den embryonale udvikling af de fleste leddyr åbner rudimentære coelomiske hulrum sig i mesodermen af i det mindste nogle somitter. Disse er snart udslettet eller bliver sammenhængende med rummet mellem de udviklende mesodermale og ektodermale strukturer og æggeblommen. Dette rum bliver til hemocoel og er således ikke beklædt af et mesodermisk peritoneum. Hos krebsdyr er de eneste coelomiske rum, der er tilbage, udskillelsesorganernes endesække og rummet omkring gonaderne.
Muskulært system: De fleste krebsdyr har en betydelig del af deres krop udgøres af tværstribede muskler. Musklerne er normalt anbragt i antagonistiske grupper: bøjere, som trækker en del ind mod kroppen, og strækkere, som strækker den udad. En flodkrebs’ bagkrop har kraftige bøjere (Figur 19-5), som bruges, når dyret svømmer bagud – dets bedste flugtvej. Stærke muskler på begge sider af maven styrer underkæben.
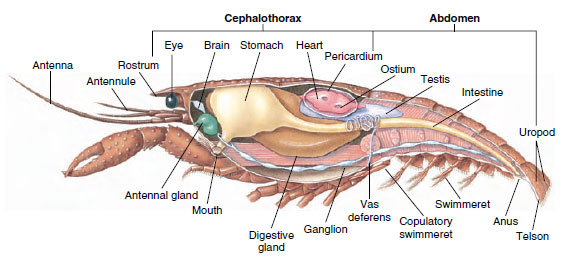
Figur 19-5
Intern opbygning af en hanekrebs.
Respiratorisk system: Åndedrætsgasudvekslingen hos mindre krebsdyr sker over tyndere områder af kutikula (f.eks. i benene) eller over hele kroppen, og specialiserede strukturer kan være fraværende. Større krebsdyr har gæller, som er delikate, fjerlignende fremspring med meget tynd kutikula. Hos decapoder omslutter siderne af carapaciet gællerummet, som er åbent fortil og ventralt (Figur 19-6). Gællerne kan stikke ud fra pleuravæggen ind i gællerummet, fra brystbenenes led med kroppen eller fra brystbenene eller fra brystskålene. De to sidstnævnte typer er typiske for flodkrebs. “Baileren”, der er en del af den anden overkæbe, trækker vand over gælletrådene, ind i gællerummet ved basen af benene og ud af gællerummet ved den forreste.
Cirkulationssystemet: Krebsdyr og andre leddyr har et “åbent” eller lacunært kredsløbssystem. Det betyder, at der ikke er nogen vener og ingen adskillelse af blodet fra interstitiel væske, som det er tilfældet hos dyr med lukkede systemer. Hæmolymphe (blod) forlader hjertet via arterier, cirkulerer gennem hemocoelen og returnerer til venøse sinusser eller rum i stedet for vener, før det kommer tilbage til hjertet igen. Annelider har et lukket system, ligesom hvirveldyr.
Et dorsalhjerte er det vigtigste fremdriftsorgan. Det er en enkeltkammeret sæk af tværstribede muskler. Hæmolymfe kommer ind i hjertet fra den omgivende sinus pericardialis gennem parvise ostier med ventiler, der forhindrer tilbagestrømning ind i sinus (Figur 19-6). Fra hjertet kommer hæmolymphen ind i en eller flere arterier. Ventiler i arterierne forhindrer tilbagestrømning af hæmolymphe. Små arterier tømmes ud i vævssinus, som igen ofte munder ud i en stor sinus sternal (Figur 19-6).
Herfra fører afferente sinuskanaler hæmolymphe til gællerne, hvis de er til stede, til udveksling af ilt og kuldioxid. Hæmolymphen vender derefter tilbage til sinus pericardialis via efferente kanaler (Figur 19-6).
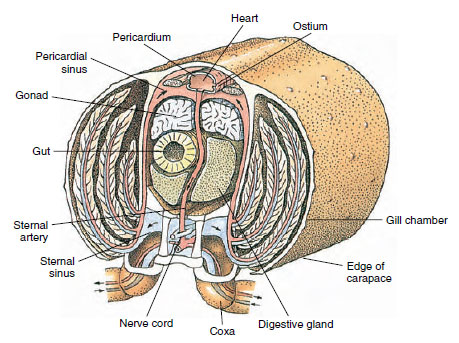
Figur 19-6
Diagrammatisk tværsnit gennem hjerteregionen af en flodkrebs, der viser retningen af blodstrømmen i dette “åbne”
blodsystem. Hjertet pumper blod til kroppens væv gennem arterier, som tømmes ud i vævets sinusser. Det tilbagevendende
blod kommer ind i sinus sternal, går derefter gennem gæller til gasudveksling og til sidst tilbage til sinus pericardialis via efferente
kanaler. Bemærk fravær af vener.
Hæmolymphe hos leddyr er stort set farveløs. Den omfatter ameboide celler af mindst to typer. Hæmocyanin, et kobberholdigt respirationspigment, eller hæmoglobin, et jernholdigt pigment, kan være med i opløsning. Hæmolymphe har den egenskab, at den kan størkne, hvilket forhindrer tab af den ved mindre skader. Nogle amøboide celler frigiver et trombinlignende koaguleringsmiddel, der udfælder koaguleringen.
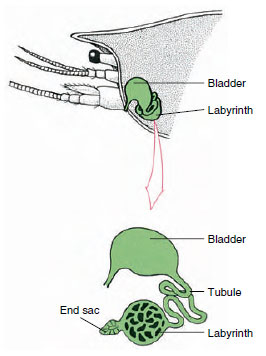
Figur 19-7
Skema af antennekirtel (grøn kirtel) hos
krebs. (I naturlig stilling er organet meget
foldet.) Nogle krebsdyr mangler en labyrint,
og udskillelsesrøret (nephridialkanalen) er
et meget oprullet rør.
Udskillelsessystemet: Udskillelsesorganer hos voksne krebsdyr er et par rørformede strukturer, der er placeret i den ventrale del af hovedet foran spiserøret (Figur 19-5). De kaldes antennekirtler eller kæbekirtler, alt efter om de åbner sig ved basis af antennerne eller ved basis af den anden kæbekrop. Nogle få voksne krebsdyr har begge dele. Udskillelsesorganer hos decapoder er antennekirtler, som i denne gruppe også kaldes grønne kirtler. Krebsdyr har ikke malpighiske rør, som er de udskillelsesorganer, der findes hos edderkopper og insekter.
Antennalkirtlens endesæk, der stammer fra et embryonalt coelomisk rum, består af en lille vesikel (saccule) og en svampet masse, der kaldes en labyrint. Labyrinten er via et udskillelsesrør forbundet med en dorsal blære, som åbner sig til det ydre ved en pore på den ventrale overflade af det basale antennesegment (Figur 19-7). Det hydrostatiske tryk i hemocoelen giver kraft til filtrering af væske ind i endesækken. Når filtratet passerer gennem udskillelsestubuli og blæren, modificeres det ved resorption af salte, aminosyrer, glukose og noget vand og udskilles til sidst som urin.
Udskillelse af kvælstofholdigt affald (hovedsagelig ammoniak) sker ved diffusion gennem tynde områder af kutikula, især gællerne, og de såkaldte udskillelsesorganer fungerer hovedsagelig til at regulere den ioniske og osmotiske sammensætning af kropsvæsker. Ferskvandskrebsdyr som f.eks. krebsfisk er konstant truet af overfortynding af deres blod med vand, som diffunderer gennem gæller og andre vandgennemtrængelige overflader. De grønne kirtler, der danner en fortyndet, saltfattig urin, fungerer som en effektiv “oversvømmelseskontrol”. Noget Na+ og Cl- går tabt i urinen, men dette tab kompenseres af gællernes aktive optagelse af opløst salt. Hos marine krebsdyr, f.eks. hummere og krabber, fungerer nyrerne til at justere saltsammensætningen i hæmolymphen ved selektiv ændring af saltindholdet i rørurinen. Hos disse former forbliver urinen isosmotisk i forhold til blodet.
Nervesystemet og det sensoriske system: Nervesystemerne hos krebsdyr og annelider har meget til fælles, selv om krebsdyrenes nervesystemer har flere fusioner af ganglier (figur 19-5). Hjernen er et par supraesophageale ganglier, der forsyner nerverne til øjnene og to par antenner. Den er forbundet med forbindelsesled til det subesophageale ganglie, en fusion af mindst fem par ganglier, der forsyner munden, vedhæng, spiserøret og antennekirtlerne med nerver. Den dobbelte ventrale nervestreng har et par ganglier til hver somit og nerver, der forsyner vedhæng, muskler og andre dele.
Ud over dette centrale system kan der være et sympatisk nervesystem, der er forbundet med fordøjelseskanalen.
Krustdyr har bedre udviklede sanseorganer end annelider. De største sanseorganer hos krebsdyrene er øjne og statocyster. De er bredt fordelt over kroppen og har taktile hår, fine fremspring af kutikula, som er særligt talrige på chelae, munddele og telson. De kemiske smags- og lugtesanser findes i hårene på antenner, munddele og andre steder.
En sæklignende statocyst, der åbner sig til overfladen gennem en dorsal pore, findes på det basale segment af hver af krebsens første antenner. Statocysten indeholder en kam, som bærer sansehår, der er dannet af den kitinagtige foring og sandkorn, der tjener som statolitter. Når dyret ændrer sin position, sendes de tilsvarende ændringer i positionen af kornene på sansehårene som stimuli til hjernen, og dyret kan tilpasse sig i overensstemmelse hermed. Ved hver skiftning (ecdysis) af kutiklen tabes statocystens kutikulære foring og dermed sandkornene. Nye korn opsamles gennem den dorsale pore efter ecdysen.
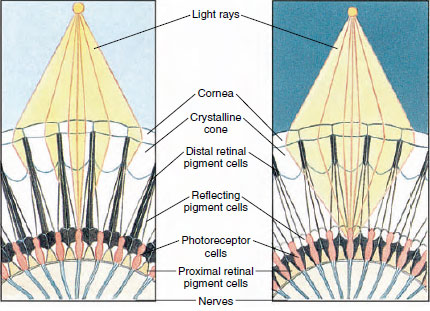
Figur 19-8
Afsnit af sammensat øje hos en leddyr, der viser migration af pigment
i ommatidia til dag- og nattesyn. Fem ommatidia er repræsenteret i
hver figur. Om dagen er hvert ommatidium omgivet af en mørk
pigmentkrave, således at hvert ommatidium kun stimuleres af lysstråler
der trænger ind i dets egen hornhinde (mosaiksyn); om natten danner pigmentet
ufuldstændige kraver, og lysstråler kan sprede sig til tilstødende ommatidia
(kontinuerligt billede, eller superpositionsbillede).
Ojnene hos mange krebsdyr er sammensatte og består af mange fotoreceptorenheder kaldet ommatidia (Figur 19-8). Den afrundede overflade af hvert øje er dækket af et gennemsigtigt område af kutikula, cornea, som er opdelt i mange små firkanter eller sekskanter, der kaldes facetter. Disse facetter er de ydre ender af ommatidia. Hvert ommatidium opfører sig som et lille øje og indeholder flere slags celler, der er anbragt i en søjleform (Figur 19-8). Der findes sorte pigmentceller mellem tilstødende ommatidia.
Bevægelsen af pigmentet i et leddyrs sammensat øje gør det muligt for det at tilpasse sig forskellige lysmængder. Der er tre sæt pigmentceller i hvert ommatidium: distale retinale, proximale retinale og reflekterende; disse er anbragt således, at de kan danne en mere eller mindre komplet krave eller muffe omkring hvert ommatidium. Ved stærk lys- eller dagtilpasning bevæger det distale retinale pigment sig indad og møder det udadgående proximale retinale pigment, således at der dannes et komplet pigmenthylster omkring ommatidium (Figur 19-8). I denne tilstand vil kun stråler, der rammer hornhinden direkte, nå fotoreceptorcellerne (retinularcellerne), for hvert ommatidium er afskærmet fra de andre. Hvert ommatidium vil således kun se et begrænset område af synsfeltet (et mosaik- eller appositionsbillede). I svagt lys adskilles distale og proximale pigmenter, således at lysstrålerne ved hjælp af reflekterende pigmentceller har mulighed for at sprede sig til de tilstødende ommatidia og danne et sammenhængende eller superpositionsbillede. Denne anden type syn er mindre præcis, men udnytter den begrænsede mængde lys, der modtages, maksimalt.
Reproduktion, livscyklus og endokrin funktion
De fleste krebsdyr har adskilte køn, og der findes forskellige specialiseringer for parring blandt forskellige grupper. Snegle er enæggede, men praktiserer generelt krydsbefrugtning. Hos nogle ostracoder er hanner sjældne, og reproduktionen er normalt parthenogenetisk. De fleste krebsdyr udrugter deres æg på en eller anden måde: grenlopper og strandsnegle har særlige ynglekamre, copepoder har ynglesække, der er fastgjort til siderne af bugen (se figur 19-19), og mange malacostracere bærer æg og unger fastgjort til deres bugtilbehør.
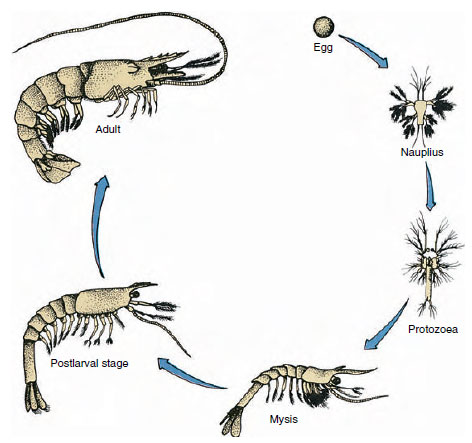
Figur 19-9
Livscyklus hos en Gulf-reje Penaeus. Penaeider gyder på 40 til 90 meters dybde. De unge larveformer er
planktoniske og bevæger sig ind mod kysten til vand med lavere saltholdighed for at udvikle sig som ungfisk. Ældre rejer vender tilbage til dybere vand
på land.
Krebsfisk har en direkte udvikling: der er ingen larveform. En lille ungfisk med samme form som den voksne og et komplet sæt vedhæng og somitter udklækkes af ægget. Derimod er udviklingen indirekte hos de fleste krebsdyr, og en larve, der i struktur og udseende er helt forskellig fra den voksne, klækkes fra ægget. Forandringen fra larve til voksen er i sidste ende en metamorfose. Den primitive og mest udbredte larve hos krebsdyrene er nauplien (figur 19-9 og 19-23). Nauplier har kun tre par vedhæng: en par ensartede første antenneled, to par ensartede antenner og to par ensartede mandibler. Alle fungerer som svømmende vedhæng på dette stadium.
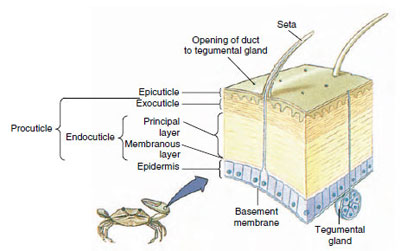
Figur 19-10
Krebsdyrs kutikulestruktur.
Den efterfølgende udvikling kan indebære en gradvis ændring til den voksne kropsform, og vedhæng og somitter tilføjes gennem en række skift, eller overtagelse af den voksne form kan indebære mere pludselige ændringer. For eksempel forløber metamorfosen hos en havtaske fra en fritsvømmende nauplius til en larve med et toskallet panser, kaldet en cyprid, og til sidst til en fastsiddende voksen med kalkplader.
Ecdyse: Ecdysis (ek´duh-sis) (gr. ekdyein, at fjerne), eller muldning, er nødvendig for at kroppen kan vokse i størrelse, fordi det ydre skelet ikke lever og ikke vokser i takt med, at dyret vokser. En stor del af et krebsdyrs funktion, herunder dets reproduktion, adfærd og mange metaboliske processer, påvirkes direkte af den fysiologi, der ligger til grund for skiftningscyklussen.
Hudskindet, som udskilles af den underliggende epidermis, har flere lag (figur 19-10). Det yderste er epicuticle, et meget tyndt lag af lipidimprægneret protein. Hovedparten af kutikula er de flere lag af procuticle: (1) exocuticle, som ligger lige under epicuticle og indeholder protein, calciumsalte og chitin; (2) endocuticle, som selv består af (3) et hovedlag, som indeholder mere chitin og mindre protein og er stærkt forkalkede, og (4) et ikke forkalkede membranlag, et relativt tyndt lag af chitin og protein.
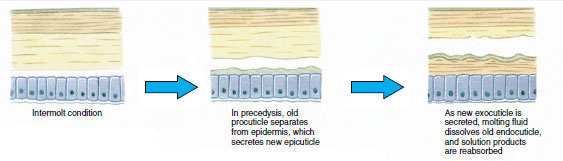
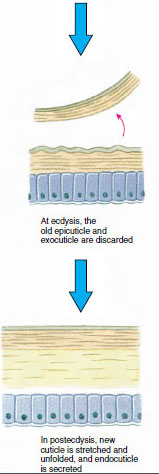
Et stykke tid før den egentlige ecdyse udvider epidermalcellerne sig betydeligt. De adskiller sig fra det membranøse lag, udskiller en ny epicuticle og begynder at udskille en ny exocuticle (Figur 19-11). Enzymer frigives i området over den nye epicuticle. Disse enzymer begynder at opløse den gamle endocuticle, og de opløselige produkter bliver resorberet og lagret i krebsdyrets krop. Nogle kalciumsalte lagres som gastrolitter (mineralske aflejringer) i mavesækens vægge. Til sidst er der kun exocuticle og epicuticle tilbage af den gamle kutikula, som underlejres af ny epicuticle og ny exocuticle. Dyret sluger vand, som det optager gennem tarmen, og dets blodvolumen øges kraftigt. Det indre tryk får kutikulaen til at flække, og dyret trækker sig ud af sit gamle exoskelet (Figur 19-12). Derefter følger en strækning af den stadig bløde nye kutikula, aflejring af den nye endokutikula, genaflejring af de bjærgede uorganiske salte og andre bestanddele og hærdning af den nye kutikula. I den periode, hvor skiftet finder sted, er dyret forsvarsløst og holder sig skjult.
 |
|
 |
 |
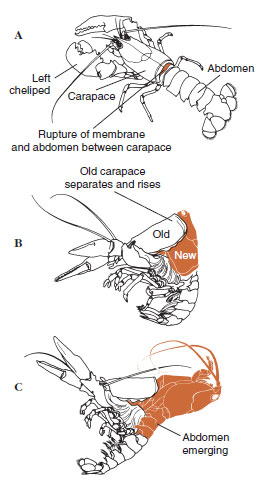
| Figur 19-12 Moltningssekvens hos en hummer, Homarus americanus. A, Membranen mellem carapace og abdomen brister, og carapace begynder langsomt at hæve sig. Dette trin kan tage op til 2 timer. B og C: Hoved, thorax og til sidst maven trækker sig tilbage. Denne proces tager normalt ikke mere end 15 minutter. Umiddelbart efter ecdysen er chelipederne udtørrede og kroppen er meget blød. Hummeren fortsætter den hurtige optagelse af vand, således at kroppen inden for 12 timer vokser ca. 20 % i længde og 50 % i vægt. Vævsvand vil blive erstattet af protein i de efterfølgende uger. |
Figur 19-11 Figur 19-11 Sekretion og resorption af hudceller under ekdyse. |
Når et krebsdyr er ungt, skal ekdyse forekomme hyppigt for at muliggøre vækst, og muldningscyklusen er relativt kort. Efterhånden som dyret nærmer sig sin modenhed, bliver intervaltperioderne gradvist længere, og hos nogle arter ophører forældningen helt og holdent. I intermoltperioderne sker der en forøgelse af vævsmassen, da levende væv erstatter vand.
Hormonel styring af ekdysecyklussen
Og selv om ekdyse er hormonelt styret, bliver cyklussen ofte igangsat af en miljømæssig stimulus, der opfattes af centralnervesystemet. Sådanne stimuli kan omfatte temperatur, dagslængde og fugtighed (i tilfælde af landkrabber). Signalet fra centralnervesystemet nedsætter X-organets produktion af et hormon, der hæmmer hudafvikling. X-organet er en gruppe af neurosekretoriske celler i hjernens medulla terminalis. Hos flodkrebs og andre tiarmede krebsdyr findes medulla terminalis i øjenstilken. Hormonet transporteres i X-organets axoner til sinuskirtlen (som i sig selv sandsynligvis ikke har en kirtelfunktion), også i øjenstilken, hvor det frigives i hæmolymphen.
Et fald i niveauet af skimmelinhiberende hormon fremmer frigivelsen af et skimmelhormon fra Y-organerne. Y-organerne ligger under epidermis nær underkæbenes adduktormuskler, og de er homol-ogus med insekternes prothorakale kirtler, som producerer hormonet ecdyson. Skimmelhormonets virkning er at igangsætte processer, der fører til ecdyse (proecdyse). Når først cyklussen er indledt, fortsætter den automatisk uden yderligere virkning af hormoner fra hverken X- eller Y-organer.
Andre endokrine funktioner: Ikke alene fremskynder fjernelsen af øjenstænglerne muldningen, men man fandt også for over 100 år siden, at krebsdyr, hvis øjenstængler er blevet fjernet, ikke længere kan tilpasse kroppens farvetegning til baggrundsforholdene. For over 50 år siden blev det opdaget, at fejlen ikke skyldes tab af synet, men tab af hormoner i øjenstænglen. Krebsdyrs kropsfarve er i vid udstrækning et resultat af pigmenter i særlige forgrenede celler (chromatophorer) i epidermis.
Koncentration af pigmentkorn i midten af cellerne giver en lysere effekt, og spredning af pigmentet i alle cellerne giver en mørkere effekt. Pigmentadfærd styres af hormoner fra neurosekretoriske celler i øjenstilken, ligesom vandring af nethindepigment til lys- og mørketilpasning i øjnene (Figur 19-8).
Udskillelse af neurosekretorisk materiale fra perikardieorganerne i perikardievæggen forårsager en stigning i hjerteslagets frekvens og amplitude.
Androgene kirtler, der først blev fundet hos en amfipode (Orchestia, en almindelig strandhopper), forekommer hos mandlige malacostracere. I modsætning til de fleste andre endokrine organer hos krebsdyr er disse ikke neurosekretoriske organer. Deres sekretion stimulerer udtrykket af mandlige kønskarakteristika. Unge malacostracere har rudimentære androgene kirtler, men hos hunnerne udvikles disse kirtler ikke. Hvis de kunstigt implanteres hos en hun, omdannes hendes æggestokke til testikler og begynder at producere sædceller, og hendes vedhæng begynder at antage mandlige kendetegn ved den næste hudskifte. Hos isopoder findes de androgene kirtler i testiklerne; hos alle andre malacostracere befinder de sig mellem musklerne i coxopoderne på de sidste brystben og er delvist fastgjort i nærheden af enderne af vasa deferentia. Selv om hunnerne ikke har organer, der ligner androgene kirtler, producerer deres æggestokke et eller to hormoner, der påvirker de sekundære kønskarakteristika.
Hormoner, der påvirker andre kropsprocesser hos krebsdyr, kan være til stede, og der er tegn på, at et neurosekretorisk stof, der produceres i øjenstilken, regulerer blodsukkerniveauet.
Fødevaner
Fødevaner og tilpasninger til fødeindtagelse varierer meget blandt krebsdyr. Mange former kan skifte fra en type fødeindtagelse til en anden afhængig af miljøet og fødetilgængeligheden, men alle bruger det samme grundlæggende sæt munddele. Mandibler og maxillae fungerer til at indtage føde, mens maxillipeds holder og knuser føde. Hos rovdyr tjener de gående ben, især chelipæderne, til at fange føde.
Mange krebsdyr, både store og små, er rovdyr, og nogle har interessante tilpasninger til at dræbe byttet. En rejeagtig form, Lygiosquilla, har på et af sine gangben et specialiseret finger, der kan trækkes ind i en rille og pludselig frigives for at gennembore et forbipasserende bytte. Pistolrejer (Alpheus spp.) har en enormt forstørret chela, der kan spændes som hammeren på en pistol og knækkes med en kraft, der lammer byttet.
Suspensionsrejsernes føde spænder fra plankton og detritus til bakterier. Rovdyrene spiser larver, orme, krebsdyr, snegle og fisk. Aasædere spiser døde dyre- og plantematerialer. Suspensionsædere, som f.eks. fe-rejer, vandlopper og strandskaller, bruger deres ben, som er forsynet med en tyk kant af setae, til at skabe vandstrømme, der fejer fødepartikler gennem setae’erne. Slamrejer (Upogebia spp.) bruger lange setae på de to første par af deres thorakale vedhæng til at si fødevaremateriale fra vandet, der cirkulerer gennem deres hule ved bevægelser af deres svømmeblodder.
Krebsfisk har en todelt mave (Figur 19-13). Den første del indeholder en mavekværn, hvor føde, der allerede er revet op af mandiblerne, kan findeles yderligere af tre kalkholdige tænder til partikler, der er fine nok til at passere gennem et setosefilter i den anden del; fødepartiklerne passerer derefter ind i tarmen til kemisk fordøjelse.
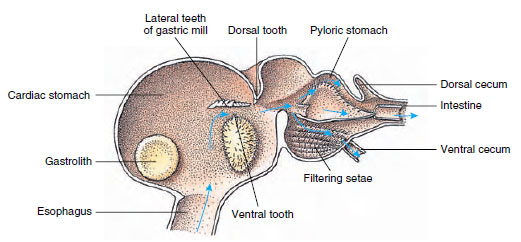
Figur 19-13
Malacostrakisk mave, der viser gastrisk “mølle” og retninger for fødebevægelser. Møllen har chitinøse ribber eller
tænder til mastikulation og setae til at si fødevarerne, før de passerer ind i pylorusmaven.
Skriv et svar